EVOLUTIONÄRE KONTINUITÄT UND DISKONTINUITÄT
Der Grundgedanke der fast vollständigen Kontinuität in der Evolution war und ist bis heute im Darwinismus so ausgeprägt, dass alle eventuellen und realen größeren Diskontinuitäten in der Geologie und Biologie als ephemäre Randerscheinungen gelten und damit für alle wesentlichen Evolutionsfragen als völlig bedeutungslos eingestuft werden. ["...metaphysical uniformitarianism is part and parcel of pure neo-Darwinism, and one of its severe weaknesses." S.N. Salthe (1987): On the trail of the unknown biology (Buchbesprechung). J. Hered 78, 213-214]. De Vries, Bateson, Johannsen und andere Pioniere der Genetik haben diese Geisteshaltung mit dafür verantwortlich gemacht, dass die Mendelschen Entdeckungen zur Existenz und Rekombination diskreter Erbfaktoren erst nach einer Verzögerung von 34 Jahren eine weitere Verbreitung und Anerkennung gefunden haben. Ja diese Pioniere selbst wurden in teilweise ungeheuer polemische Auseinandersetzungen mit ihren darwinistischen Zeitgenossen verwickelt, welch letztere alles daransetzten, den 'Mendelismus' und die damit verbundenen Diskontinuitätsansätze zum Ursprung der Arten zu unterbinden.
Mayrs Kommentar zu dieser Problematik lautet u.a. 1982, pp. 724/725:
For a long time after 1900 it was widely believed that continous variation obeyed entirely different laws of inheritance than those of Mendel, and this might add another reason for the neglect of Mendel's work. After all, gradual continous variation was widely considered after 1859 as the only variation of interest to these evolutionists.
It took many years (after 1900) before the "true Darwinians", as they liked to call themselves, understood that gradual evolution and continous variation could be explained in Mendelian terms.
Zu diesen wahren Darwinisten gehörten bis in die 30er Jahre unseres Jahrhunderts hinein Biologen wie z.B. Rensch und Mayr selbst (Mayr 1982, p. 554).
Diese Auffassung der evolutionären Kontinuität - seit der 'evolutionären Synthese' in Verbindung mit der Mendelschen quantitativen Vererbung - ist so tief verwurzelt, dass sie bis heute als wesentliche Grundvoraussetzung und -anschauung des Neodarwinismus zu bezeichnen ist. Dieser Punkt klingt in den obigen Zitaten von Mayr, Fidley und Lorenzen schon an und wird in der gesamten neodarwinistischen Literatur immer wieder betont. "In due time it was realized that the spectacular De Vriesian mutations were exceptional phenomena and that the normal genetic changes are "small" mutations (Baur, East, Johannsen, Morgan) which...have only slight or even invisible effects on the phenotype" (Mayr 1970, p. 169 und ähnlich 1979, pp. 40/41; ebenso Heberer 1959, Stebbins 1974, Dobzhansky et al. 1977, Rensch 1972, 1977, I. R. Bock 1984 u.v.a.). Diese Mutationen sind es, die mit ihren schwachen oder sogar unsichtbaren Wirkungen auf den Phänotyp und mit ihrer relativ niedrigen Frequenz die kontinuierliche Evolution garantieren.
Nilsson hat die Geschichte des Mutationismus im Kapitel III seiner großen Arbeit von 1953 ausführlich geschildert. "Der heutige Stand der Theorie" - so die Überschrift zum Abschnitt 4 des Kapitels - könnte die Beurteilung aus neo-darwinistischer Sicht von heute sein. Wir lesen beispielsweise p. 161:
Die Auffassung der Genetiker über die Wertlosigkeit der Großmutanten ist auch heute fast allgemein [akzeptiert]. Hiermit hat man auch ganz definitiv die Mutationslehre von HUGO DE VRIES, nach der vor allem die großen Sprünge, die diskontinuierliche Variabilität, von fundamentaler Bedeutung waren, aufgegeben. Hier lag indessen gerade die Stärke der neuen Auffassung der Variabilität. Denn hier wurde die Evolution aus dem Schneckentempo befreit, die die ganze Erscheinung unwahrscheinlich zu machen gedroht hatte. Die Evolution als eine Mutation und die Mutation als Saltation, ist in der heutigen Genetik ein überwundenes Stadium.
Um so intensiver klammert sich die heutige Artbildungsforschung an die alte DARWINsche Lehre von der Summierung ganz unbedeutender quantitativer Differenzen als Ursache der Entwicklung (Hervorhebung im Schriftbild im Original gesperrt.)
Für die Genetiker allgemein kann man das heutzutage aber nicht mehr behaupten. Zumindest bei den Transposongenetikern werden "complex changes in a single event" wieder ernsthaft für die Evolutionsfrage diskutiert. Für die Synthetische Evolutionstheorie waren jedoch schon in den 40er Jahren die 'antidarwinistischen Theorien der Saltation und genetischen Eigengesetzlichkeit' (vgl. p. 438) so endgültig widerlegt, dass sich die Biologen im großen und ganzen auf die Theorie der kontinuierlichen Entwicklung durch Mikromutationen und Selektion einigen konnten. Mayr meint 1979, p. 20:
Niemand, der nicht gründlich über die jüngsten Fortschritte in der Genetik und der Evolutionslehre unterrichtet ist, kann wirklich ganz ermessen, wie stark die Position der Darwinschen Evolutionstheorie ist!
(p. 84:) Es ist heute unbestritten, daß es keine saltatorische Evolution gibt...
Die verschiedenen neueren Ansätze zur Evolutionstheorie wie Neutralismus, Punktualismus und Transposon-verursachte komplexe Veränderungen in einem einzigen Schritt liegen zwar alle noch im Rahmen einer naturwissenschaftlichen Interpretation der Frage nach dem Ursprung der Organismenwelt (in Sinne einer Rückführung auf physiko-chemische Gesetzmäßigkeiten), sie haben jedoch den neo-darwinistischen Rahmen längst gesprengt, indem sie sowohl die Entwicklungsgeschwindigkeit als auch die herkömmliche Mutations- und Selektionstheorie derart modifiziert bzw. aufgegeben haben, dass führende Neodarwinisten selbst solche Ansätze als 'antidarwinistisch' einstufen.
Hat jedoch Mayr mit seiner peripatrischen Speziationstheorie nicht auch eine beschleunigte Evolution der Arten vertreten, an die sogar der Punktualismus anschließen konnte (vgl. p. 83)? Der Punktualismus steht nach Mayrs Auffassung "keineswegs im Widerspruch zu Darwin - allerdings mit dem Vorbehalt, daß es sich immer um eine (wenn auch sehr schnelle) Änderung von Populationen handelt. Und jede Populations-Evolution ist, definitionsgemäß, eine allmähliche, also Darwinsche Evolution" (Mayr 1984). Merke: Sehr schnell heißt auf darwinistisch immer noch allmählich. Zum Fossilbericht schreibt derselbe Autor 1984:
Sprünge, die die Fossilien-Befunde vorspiegeln, sind ohne Zweifel ein Artefakt der Lückenhaftigkeit der Fossil-Befunde. Unter den Fossilienfunden aufzutreten ist nur für weitverbreitete, äußerst individuenreiche Arten wahrscheinlich. Überreste von den individuenarmen, eng lokalisierten Gründer-Populationen werden wohl nie als Fossilien gefunden werden. Dieser Negativ-Befund berechtigt aber nicht, Sprünge zu postulieren.
Was die allmähliche Evolution im konkreten Falle in der Paläontologie bedeuten kann, veranschaulicht uns der Autor am Beispiel der Zuwachsrate der Zahnlängen in der Pferdeevolution und der 'Selektionsdrucke im Gleichgewicht' 1967, p. 193: "...tatsächlich betrug das Ausmaß ihrer Zunahme nur etwa 1 mm in einer Million Jahren (Simpson 1944)."
Vgl. zu dieser Kontinuitätsidee, die allein schon durch die Variabilität der Zahnlängen heutiger Pferde ad absurdum geführt wird, die Ausführungen pp. 314-324.
Ich möchte an dieser Stelle noch einmal darauf hinweisen, dass es ja gerade das Anliegen des Punktualismus ist, statt der Erklärung von der allgemeinen Lückenhaftigkeit der paläontologischen Befunde 'die bekannte fossile Überlieferung für bare Münze zu nehmen' (Simpson 1984, vgl. Zitat p. 321). Wenn die Evolution im Rahmen individuenarmer, eng lokalisierter Gründer-Populationen einer Darwinschen allmählichen Evolution folgt und die Population sich schließlich weit verbreitet, müssten wir zumindest genauso häufig Zwischenformen zwischen den Familien, Ordnungen und Klassen etc. finden wie innerhalb solcher Gruppen; denn es muss in diesem Falle ja zur Überbrückung der Unterschiede und Klüfte auch entsprechend viele Gründerpopulationen mit allmählich weiter geographischer Verbreitung und anschließend wieder neuen Gründer-Populationen gegeben haben, um so schrittweise von einer Ordnung und Klasse zur nächsten zu gelangen. Vor dem Auftreten einer neuen Klasse müsste sich dieser Prozess Hunderte und Tausende von Malen wiederholen und entsprechend müssten von diesen geographisch weit verbreiteten Zwischenformen (vom ältesten Präkambrium bis zum Pleistozän) genauso viele Funde gemacht werden wie innerhalb der bekannten Familien, Ordnungen und Klassen. Das trifft jedoch nicht zu (vgl. p. 298 und p. 314 f.). Meines Wissens behauptet kein Vertreter dieser modifizierten Darwinschen Artentstehungstheorie, dass in einer kleinen Gründerpopulation schlagartig die Unterschiede zwischen Familien, Ordnungen und Klassen entstehen würden, wozu noch der Punkt Höherentwicklung erwähnt werden müsste. Weder die Erfahrungstatsachen der Genetik (einschließlich Polyploidie, Genduplikationen und Transposons) noch theoretische Postulate zur Entstehung neuer Gene und zur Höherdifferenzierung könnten eine solche Theorie glaubhaft machen.
Shute schrieb schon 1962, p. 189 zu dieser von der Darwinschen Theorie der graduellen Transformation weitverbreiteter Populationen abweichenden Speziationsbehauptungen Mayrs: "Clever as they are, they are fictional until demonstrable."
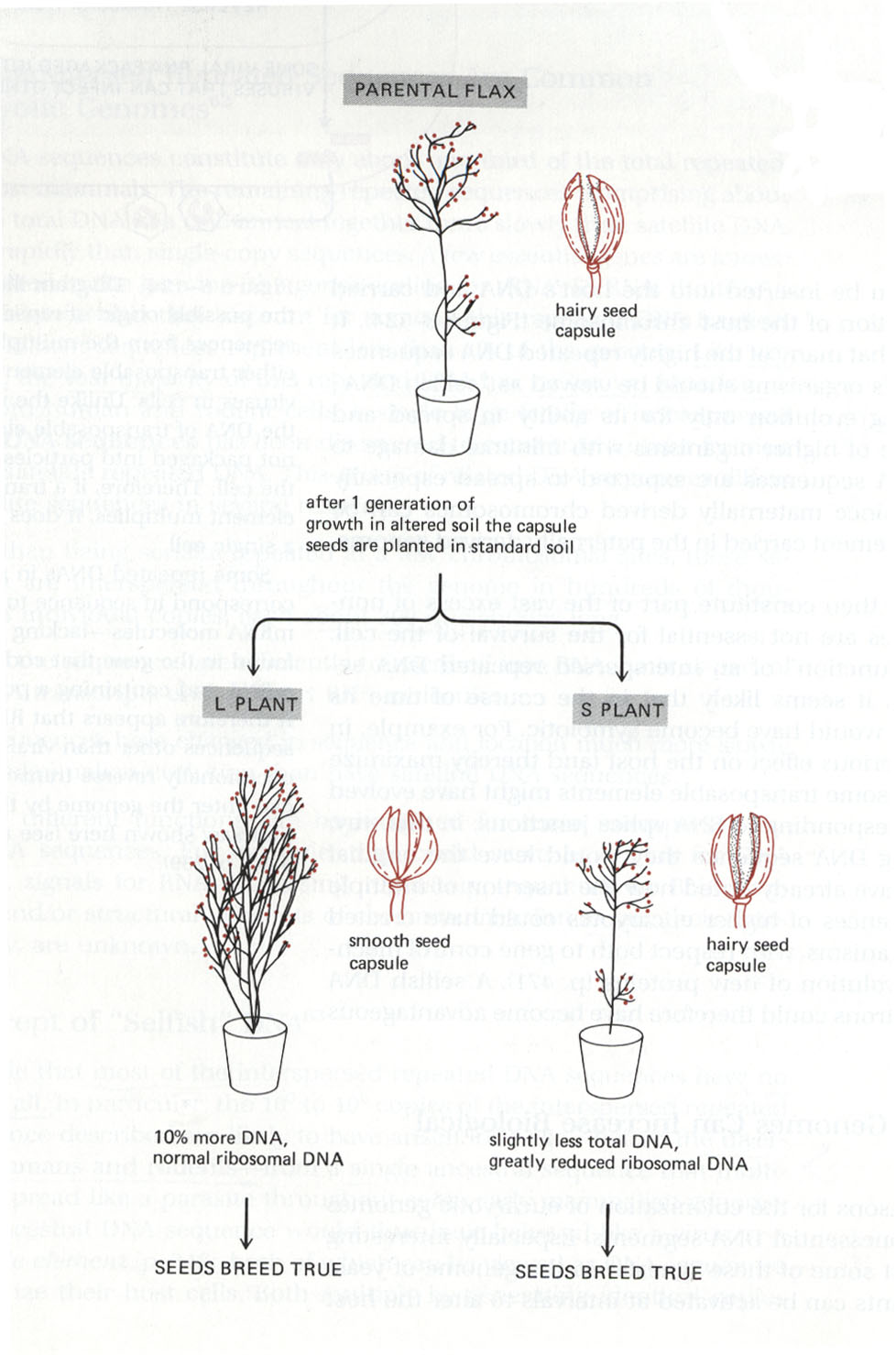
Zurück zu den Transposons: Welche Befunde sprechen nach Syvanen und anderen dafür, dass durch Transposons komplexe Veränderungen in einem einzigen Ereignis induziert werden könnten? Neben den pp. 138-l43 diskutierten Fällen wäre hier das Flachsbeispiel zu nennen. Als Antwort auf eine Reihe ungewöhnlicher Bedingungen (wie zum Beispiel zu viel Phosphate oder zu wenig Stickstoff im Boden) können Flachspflanzen Tochterpflanzen erzeugen, die sich im Wachstum der Gesamtpflanze und in der Behaarung der Samenkapseln unterscheiden. Alberts et al. berichten zusammenfassend 1983, pp. 474/475:
At least in some cases, a mixture of plant types is produced, many of which breed true and will not change further, even if soil conditions are altered. Two changes have been investigated in detail: one results in a considerably shorter plant of similar morphology (called S) and another in a plant (called L) that is very slightly shorter but has many more branches and a hairless seed capsule.
Abb. 57: Einige starke Veränderungen beim Flachs. Aus Alberts et al. 1983.
Biochemical analysis shows that major changes in the DNA have occured in the sudden transition from the parent plant to the S or L plant. Although no change in the morphology of the chromosomes is detectable, the L plant contains 10 % more DNA in each cell than does the parent because of the differential amplification of many different DNA sequences. The S plant has about 6 % less DNA per cell than the parent, with a selective deletion of certain DNA sequences. For example, the number of ribosomal RNA genes is halved.
Was sich im vorliegenden Falle genau auf der DNA-Ebene abspielt, ist bisher nicht bekannt. Cullis, der nach Beschreibung der stressinduzierten erblichen Veränderungen beim Flachs durch Durrant 1962 und 1971 das Phänomen erstmals genauer auf der DNA-Ebene untersucht hat, bemerkt in seiner Arbeit von 1985 u.a., dass nicht bei allen Linien einer Leinart erbliche Veränderungen durch die Umwelt induziert werden können, und dass die Basis für diese Unterschiede noch unbekannt ist. Weiter lesen wir auf p. 165:
The mechanisms by which these DNA rearrangements occur are also presently unknown. Possible means of achieving variation include unequal recombination, amplification by unscheduled DNA synthesis and transposition. The latter introduces one further variable not considered here, namely the role of transposable elements in DNA variation. It is known that integration and excision of transposable elements can cause amplifications and deletions, but whether they can do so on the scale described here is unclear. It is possible that they may be one of the subsets of the DNA which can be activated by stress.
Bei einigen Flachslinien unterscheidet sich die Kopienzahl der hochrepetitiven Sequenzen voneinander. Solche Unterschiede konnten auch in Kalluszellen festgestellt werden. Eine spezielle Untereinheit der 5 S Gene ist besonders von den Veränderungen betroffen (Cullis und Cleary 1985, p. 303). Nach Auffassung der Autoren kann man beim Flachsgenom mindestens zwei Bereiche unterscheiden, von denen einer schnell reguliert ("modulated") werden kann (p. 308).
Alberts et al. heben im Anschluss an das Flachsbeispiel hervor, dass analoge erbliche Veränderungen vor mehr als 30 Jahren beim Mais [Und in den letzten Jahren bei wenigstens 23 weiteren Pflanzenarten mit intraspezifischer Variation der 1 c DNA Menge zwischen 4 und 288 %, z.T. mit adaptiver Relevanz (Bennet 1985)] und mit molekulargenetischen Details auch bei Bakterien, Hefen und Drosophila festgestellt worden sind und bemerken 1983, p. 475 u.a.:
The picture that emerges is one in which either chance or severe environmental stresses change the internal environment in an organism and activate certain unknown components that cause transposable-element-like DNA sequences to move in and out of chromosomes, sometimes amplifying, deleting, or changing the pattern of expression of neighboring DNA sequences in the process. The exact changes caused by each such cataclysmic event will be determined at least partly by chance, and the level, timing, of pattern of expression of important genes in the progeny organisms will be stably altered in different ways.
It seems likely that the ability to undergo such occasional dramatic DNA rearrangements makes an important contribution to the evolution of organisms. If so, it may be in part for this reason that cells succeed so well in carrying the repeated DNA sequences that appear to be parasitizing their genomes.
Der Begriff Zufall als eine der Ursachen der Transposonaktivierung scheint beim gegenwärtigen Stand unseres Wissens mehr eine Beschreibung unserer Unkenntnis zu sein als eine mit naturwissenschaftlichen Mitteln verifizierbare Erklärung des Phänomens.
Die andere inzwischen wiederholt bestätigte Erklärung, nämlich die Aktivierung als Antwort auf besondere Stresssituationen, wobei u.a. Deletionen und Amplifikationen festzustellen sind, erinnert an die Ausführungen pp. 377 und 431 f. zur Regulation der Genamplifikation. Die Erweiterung auf Deletionen gibt einen Hinweis darauf, dass der DNA-Abbau bei spezialisierten Formen (vgl. p. 406 f.) zum Teil reguliert sein könnte. Auf der anderen Seite sind Deletionen, die durch eine Vielzahl natürlicher und künstlicher Agenzien verursacht werden, in Hunderten von Studien sicher nachgewiesen.
Beim Beispiel der möglichen Transposonaktivität beim Flachs und anderen Organismen erhält man den Eindruck, als ob im Organismus als Antwort auf die veränderten Umweltbedingungen ein genetisches Programm abläuft mit der Aufgabe, für die nächste Generation eine Anzahl neuer Genotypen zur Bewältigung der neuen Umwelt zur Verfügung zu stellen. Diejenigen Genotypen, die der neuen Situation am besten gewachsen sind, werden dann das Bestehen der Art garantieren. Den letzteren Punkt möchte ich besonders hervorheben: keine der vielen durch Transposons bedingten "kataklysmischen Ereignisse" haben auch nur einen Hinweis darauf gegeben, dass sie über die Artgrenzen (im Sinne des genetisch-plasmatischen Artbegriffs) hinweggehende genetische Veränderungen hervorrufen könnten. Beim obigen Flachsbeispiel würden vermutlich nicht einmal rein morphologisch orientierte Systematiker zwei Arten unterscheiden, wenn sie solche Formen in der Natur fänden. Dass das auch auf die pp. 138-143 diskutierten Drosophila-Beispiele zutrifft, wurde schon betont.
Es kommt nun auf die Definition des Evolutionsbegriffs an, ob man der oben zitierten Aussage folgen möchte, dass solche dramatischen DNA-Umordnungen "an important contribution to the evolution of organisms" leisten oder nicht. Für die Makroevolution gibt es dafür bislang keine Anhaltspunkte. Denkt man jedoch an die Millionen von Spezies im Sinne des morphologischen und auch des neo-darwinistischen Artbegriffs (s.o.), so kann das gut zutreffen: Wir haben ja gesehen, dass Millionen von Arten und Tausende von Gattungen, die nach diesen Artbegriffen unterschieden werden, nach dem genetisch-plasmatischen Artbegriff nichts weiter sind als Populationen verschiedener Rekombinanten weniger (Grund-) Arten. Dass die meisten spezialisierten Formen jedoch ihre ursprüngliche Anpassungsfähigkeit (ganz im Sinne der Degenerationstheorie) verloren haben, ist ein vielfach abgesichertes biologisches Faktum, das wir unter anderem an Inselpopulationen besonders eindrücklich bestätigt finden (vgl. pp. 410-419).
Dabei ist hervorzuheben, dass die "kataklysmischen" genetischen Ereignisse als autogenetische Vorgänge nicht von der behaupteten allesbeherrschenden neo-darwinistischen Selektion dirigiert werden. Erst auf organismischer Ebene kann der Faktor Selektion in der nächsten Generation wieder wirksam werden.
Aus der Bakteriengenetik stammt ein besonders eindrucksvolles Beispiel für möglicherweise Transposon-induzierte rekurrente Variation und in einem gewissen Ausmaß auch Degeneration. Heumann et al. berichten 1984 über ihre Rhizobiaceen-Studien unter anderem:
(p. 425:) All bacterial strains classified into the family Rhizobiaceae can be induced to undergo a fundamental genome arrangement. The special structure of their genome allows the formation of five distinctive phenotypes, each one adapted to a different habitat (Fig. 58).
This genome rearrangement can be induced by DNA-damaging agents, UV irradiation or chemical mutagenesis. For expression, cells have to be protected against photorepair and their replication has to be reduced by stress treatment. The rearrangement process is, with special exceptions reversible.
(p. 427:) Several strains of all six classical Rhizobium species and several Agrobacterium strains (Bergey's Manual 1974)...were subjected to the rearrangement treatment described. All of them responded with the formation of one or several of the new strains arranged into five classes listed in Table 4 and Fig. 1. Each arrangement step presented in Table 4 has been repeated many times during the last 10 years. As a result of this process only such strains can be shown that fit distinctly into our five class order, no intermediates or aberrant types are formed. Since the process of rearrangement is generally reversible as indicated by the arrows in Fig. 1, the strains of the newly defined classes III, IV and V, beside Agrobacterium = class one and Rhizobium = class II, also display the rearrangement into one or several of the other four classes. (s. Abbildung und Tabelle nächste Seite.)
Aufgrund der klassenspezifischen bakteriologischen und molekularen Charakteristika schlagen die Autoren vor, die fünf Klassen als fünf getrennte Arten zu definieren. Doch weisen die Verfasser (p. 425) darauf hin, dass sie in der vorliegenden Arbeit diese fünf Rhizobiaceen-'Arten' Klassen nennen, um eine verfrühte taxonomische Entscheidung zu vermeiden. Die Reversibilität spricht jedoch gegen ihren Vorschlag, hier eigene Spezies zu unterscheiden. Andernfalls würden wir den Speziesbegriff wieder von allen möglichen Umständen abhängig machen, unter denen sich eine 'Art' gerade bewährt oder in eine andere 'verwandelt': Was heute noch Art I wäre, wäre nach UV-Bestrahlung morgen schon Art II (aber nur bei Unterbindung der Photoreparatur und reduzierter Replikation durch Stress). Ein Teil der Spezies II verwandelt sich dann nach EMS-Behandlung in Spezies III usw., um schließlich wieder als Spezies I in Erscheinung zu treten. Die Reversibilität der Formen wird von den Autoren wiederholt betont und detailliert beschrieben. Auf der Seite 430 lesen wir z.B.:
Beside the high class specificity, there are many bacteriologic and genetic criteria that indicate the relatedness of the five classes. Most important is the reversibility of the rearrangements between the five classes. This reversibility is indicated in Fig. 1 by arrows: Any strain subjected to the treatment described will respond with rearrangements producing one or several other strains, but only such strains appear that fit into our rearrangement scheme.
Wir geben dazu die Fig. 1 und die Tabelle 4 der Autoren als Abb. 58 und Tab. 16 auf der nächsten Seite wieder. Es handelt sich damit um verschiedene Modifikationen, in denen die eine Rhizobiumart auftreten kann, und Almon und Baldwin haben schon 1933 in dieser Frage den richtigen Schluss gezogen, wenn sie berichten (zitiert nach Heumann et al. p. 433):
...six culture types markedly different from Rhizobium trifolii in its typical form have been isolated from cultures of this organism under conditions which have reduced opportunity for outside contamination to the minimum.
...If these six others are to be interpreted as different forms of one, it should be equally easy to conceive of the typical form as being related and to call the seven types of cultures different manifestations of the possibilities inherent in one species.
Da diese verschiedenen Manifestationen der einen Spezies auch mit Zustandsänderungen der DNA einhergehen (Details vgl. Heumann et al.), veranschaulichen sie die Regel der Rekurrenten Variation besonders treffend. Weiter sei hervorgehoben, dass die Autoren die in der Klasse 2 auftretende Irreversibilität mit Deletionen in Verbindung bringen (p. 435):
The data in Fig. 58 and Table 16 show that the rearrangement between all classes is reversible except for the structural alteration of the nodulating Rhizobia class IIc - a to Agrobacterium and minimal-type, class Ib and Ia. This occurs only unidirectionally: from min-type bacteria, strains of the group R. leg, R. phas, R. trif have never been obtained. Obviously this must be evaluated in connection with the plant interaction of these two classes (Fig. 58). It seems that a decreasing complexity of host interaction exists in the sequence presented and that each step is connected with a specific deletion of genetic information.
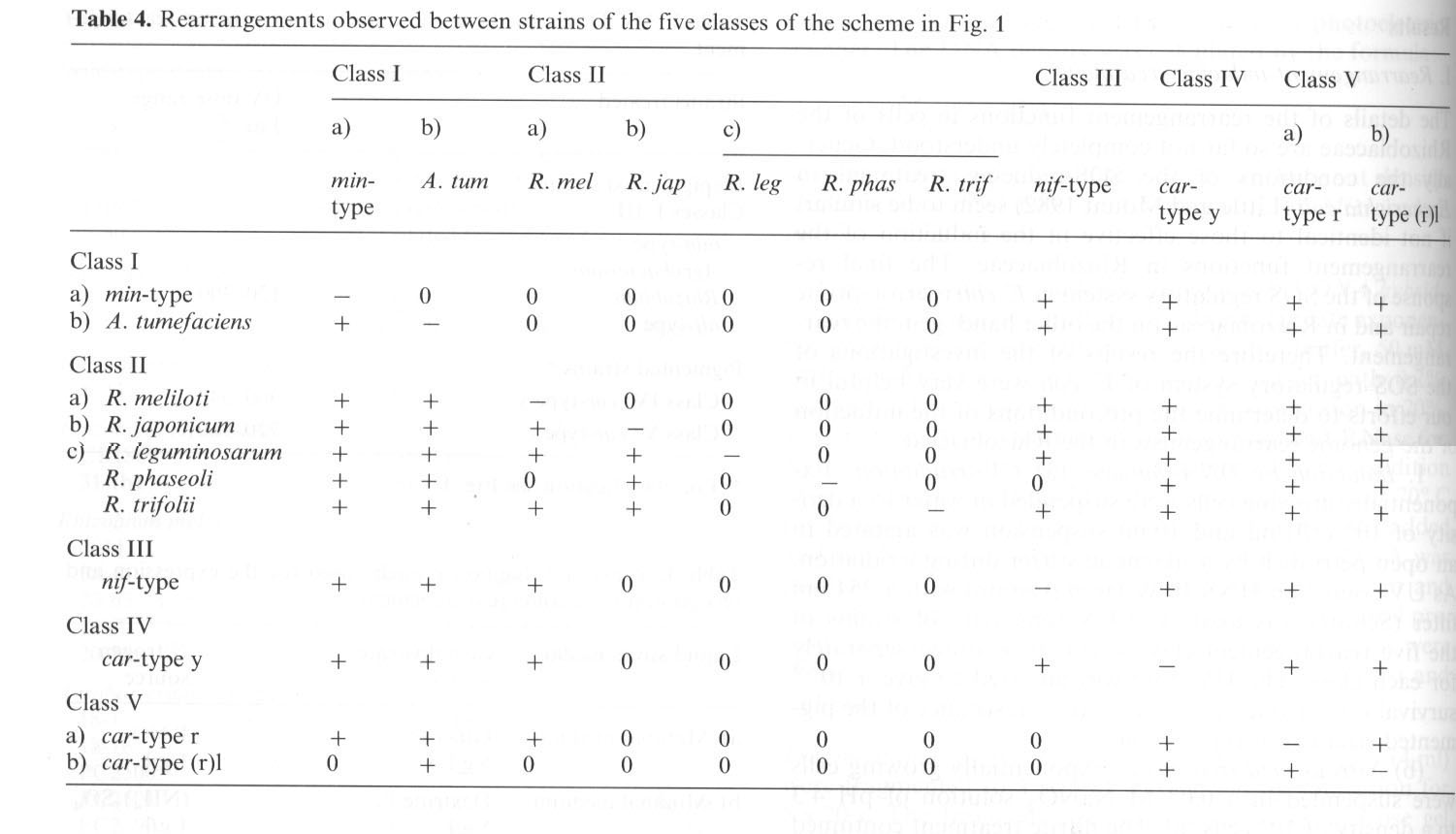
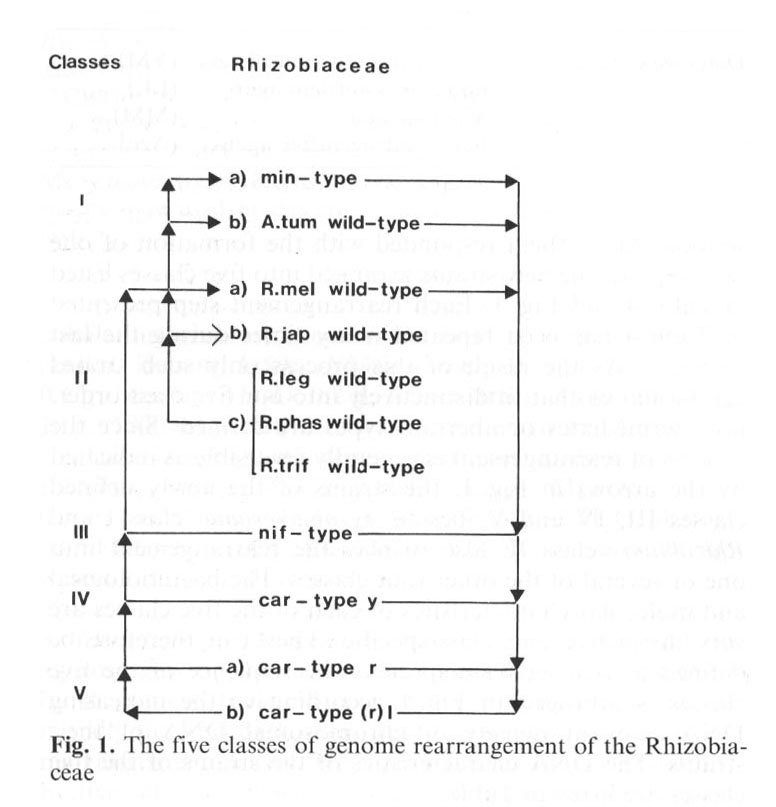
Abbildung und Tabelle aus Heumann et al. 1984.
Auf den Seiten 121 bis 265 wurde im Detail diskutiert, warum durch Degeneration bedingte (und damit sekundäre) irreversible Barrieren nicht als artentrennend aufgefasst werden können, so dass wir das Prinzip dieser Argumentation trotz der Besonderheiten der Bakteriengenetik nur auf den vorliegenden Fall übertragen brauchen.
Nach der kontinuierlichen Evolution des Neodarwinismus müssten in großen Zeiträumen durch Hunderte und Tausende von Mutationen mit geringen oder sogar unsichtbaren Wirkungen die verschiedenen Formen dieser Art von einem (unbekannten) Vorfahren herzuleiten sein, wobei die Selektion die verschiedenen Entwicklungswege völlig beherrschen würde. In der Realität treten die verschiedenen Formen in Wochen und Monaten ohne die postulierten Übergangsserien auf und sind direkt voneinander ableitbar! Das Programm der Formgestaltung liegt in den 'Arten' bereits vor und wird unter besonderen Umweltverhältnissen abgerufen. Der Faktor Selektion kann erst wieder nach den erfolgten Ereignissen wirksam werden. Bei diesen in großen Schritten wirksam werdenden komplexen Veränderungen selbst ist die Selektion ohne Bedeutung. Sie schafft weder das Programm noch dessen Expression. Diese Prozesse verlaufen nach erfolgter "Einschaltung" eigengesetzlich.