4. DIE MECHANISCHE ISOLATION
Als vierten Punkt der präzygotischen Isolationsmechanismen hat Stebbins die mechanische Isolation aufgeführt (vgl. p. 68): Die Fremdbefruchtung wird durch Unterschiede im Bau der Fortpflanzungsorgane (Genitalien der Tiere, Blüten der Pflanzen) verhindert oder eingeschränkt.
Stebbins selbst kommentiert den Punkt für die Tierwelt wie folgt (1980, p. 127):
Eine mechanische Isolation von Tieren, die auf die Unterschiede im Bau der Geschlechtsorgane zurückzuführen ist, wurde in der Vergangenheit von mehreren Zoologen als wichtige Art einer Schranke gesehen. Neuere Beobachtungen und Experimente haben jedoch gezeigt, daß diese Unterschiede relativ unwirksam in der Verhinderung von Fremdpaarung zwischen Arten sind.
Mayr fasste 1967, pp. 88/89 die Hauptpunkte zusammen (auszugsweise zitiert):
Bald nach der Entdeckung der mannigfaltigen strukturellen Unterschiede in den Genitalarmaturen verschiedener Arten von Insekten wurde von DUFOUR (1844) behauptet, daß sie eine Schloß- und Schlüssel-Funktion haben und so Bastardierungen zwischen Individuen verschiedener Arten verhindern.
Lange Zeit wurde die mechanische Isolation als ein höchst wirksamer Isolationsmechanismus betrachtet, besonders als sich herausstellte, wie weit verbreitet bei verschiedenen Ordnungen der Insekten Unterschiede im Bau der Genitalien sind.
KARL JORDAN (1905) zog jedoch aufgrund umfassender, ins einzelne gehender Untersuchungen den Schluß, daß die DUFOURsche Hypothese keine Gültigkeit besitzt. Unter 698 Sphingiden-Arten, die er analysierte, waren 48 in ihren Genitalien nicht von anderen Arten der Familie verschieden, während bei etwa 50 Prozent der Arten mit geographischer Färbungsvariabilität auch eine geographische Variabilität in den Genitalarmaturen bestand. Seitdem hat sich vieles zusätzliche Informationsmaterial angesammelt, das die geringe Bedeutung der Genitalarmaturen als Isolationsmechanismen unterstreicht (s. GOLDSCHMIDT 1940 und DOBZHANSKY 1951). So zeigte BEHEIM (1942), daß bei den Laufkäfern Bastarde sogar bei Arten mit sehr verschiedenen forceps auftreten können, zum Beispiel bei Carabus coriaceus und C. violaceus. Bei Marienkäfern der Gattung Hippodamia können Bastarde leicht zwischen Arten erzeugt werden, die betonte Unterschiede in ihren Genitalien zeigen (SHULL 1946). ROSEN und GORDON (1953) kamen zu demselben Schluß für poeciliide Fische, KULLENBERG (1947) für Hemipteren, KUNZE (1959) für Homopteren, GERING (1953) für agelenide Spinnen und EDGREN (1953) bei Schlangen. Bei einigen Insektenarten führt ausgeprägter Polymorphismus des Baues der Genitalarmaturen der Männchen nicht zu Unverträglichkeit.
In ähnlicher Weise schreibt Dobzhansky (1977, p. 174):
Dufour believed that the lock-and-key relationship of the genitalia makes species separate and unchangeable. This is undoubtedly an exaggeration. Not only large and small individuals of the same species but sometimes different species with clearly distinguishable genitalia copulate successfully. Nevertheless, it has often been observed, particularly in laboratory experiments, that interspecific copulation results in injury or even death of the participants. An example of this is copulation of Drosophila pseudoobscura females with D. melanogaster males. Females and males of dog breeds very different in size may be attracted to each other but unable to copulate.
Im Großen und Ganzen wird der mechanischen Isolation von zoologischer Seite kein großer Stellenwert mehr beigemessen. (In Klammern sei vermerkt, dass man Tatsachen und Gedankengängen, die gegen die Bedeutung eines zur Artkonstanz führenden Isolationsmechanismus sprechen, sehr aufgeschlossen ist.)
Bei höheren Pflanzen sieht die Beurteilung dieses Isolationsmechanismus anders aus. Auch scheint die in Klammern vermerkte Gefahr der Konstanz hier weniger zuzutreffen. Sowohl Grant (1963, 1976, 1981) als auch Stebbins (1980) und Briggs und Walters (1984) führen zahlreiche Beispiele für diese Isolation in ihren Arbeiten auf. Hier Stebbins' Urteil und vielleicht eines der besten Beispiele mechanischer Isolation im Pflanzenreich (p. 127):
Bei höheren Pflanzen sind die wirksamsten präzygotischen Isolationsmechanismen Unterschiede im Bau der Blüten, die entweder vollkommen unterschiedliche Bestäuber zu verschiedenen Arten locken, oder die Chancen der Kreuzung sehr vermindern, wenn ein bestimmter Bestäuber nacheinander die Blüten zweier verschiedener Arten besucht. So haben die meisten Arten von Pentstemon, dem Bartfaden, blaue bis violette Blüten, aber ihre Größe und Form variiert stark von einer Art zur anderen. Einige sind an die Bestäubung durch große Bienen, wie die Holzbiene, andere an Hummeln, noch andere an kleine solitäre Bienen oder an Wespen angepaßt. Daneben haben einige wenige Arten leuchtend rote Blüten mit Röhrenform, passend für die Schnäbel von Kolibris, ihren Hauptbestäubern.
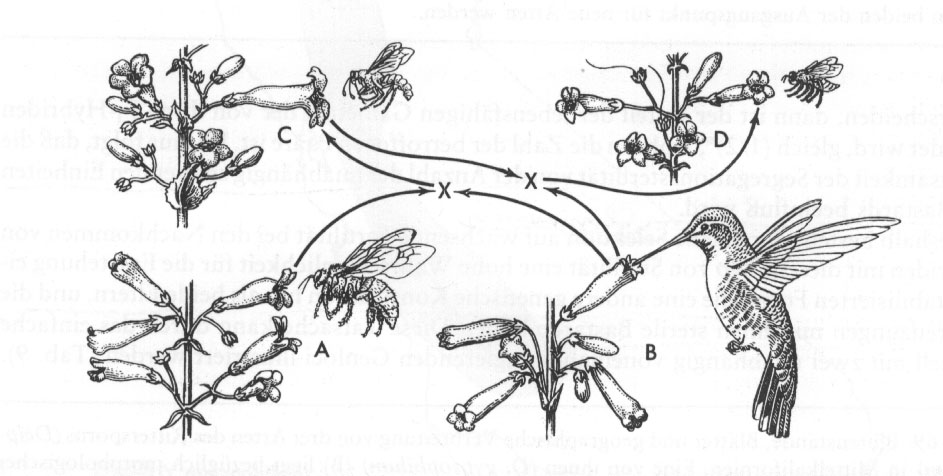
(pp. 154/155:) Bei Pflanzen können Rekombinationen der Gene, die Isolationsmechanismen steuern, neue Genotypen erzeugen, die teilweise von ihren beiden ursprünglichen Eltern isoliert sind. Dies ist sowohl in bezug auf Isolation durch Blütenbau als auch durch Sterilität bei der Segregation der Hybriden klar demonstriert worden. Für die Isolation durch Blütenbau bietet die Gattung Bartfaden (Pentstemon) aus dem Westen der USA gute Beispiele. Hierher gehört eine große Artenzahl, die sich hauptsächlich in Form, Größe und Farbe der Blüten unterscheidet. Vier der bekanntesten Arten aus Südkalifornien sind in Abb.15 gezeigt. Eine aus Kiefernwäldern im Gebirge (Bergbartfaden) hat große zweilippige, weit geöffnete Blumenkronen in leuchtendem Blau. Eine zweite, als "Roter Hornist" bekannt, hat rote röhrenförmige Blüten und ist in den trockenen Teilen Südkaliforniens weit verbreitet. Eine dritte Art, der Prachtbartfaden, kommt auf gestrüppbedeckten Hügeln in einem relativ neuen Habitat vor, das in seiner Charakteristik zwischen denen der beiden anderen Arten liegt. Der Prachtbartfaden hat blaue oder schwach violette Blüten, intermediär in der Form zwischen den beiden erstgenannten. An einigen wenigen Stellen wurden natürliche Hybriden zwischen dem Bergbartfaden und dem Roten Hornisten gefunden, die in vieler Hinsicht dem Prachtbartfaden sehr ähnlich sind. Die drei Arten werden normalerweise von den folgenden Bestäubern besucht: Der Bergbartfaden von großen Holzbienen (Xylocopa), der Rote Hornist von Kolibris und der Prachtbartfaden von solitären Wespen (Pseudomasaris).
Wir können die mögliche Folge der Vorgänge, die zur Entstehung des Prachtbartfadens führten, folgendermaßen rekonstruieren. In der Vergangenheit waren die Berge Südkaliforniens nicht von Gestrüpp bedeckt, sondern mit Ausnahme der steilen, trockenen Hänge bewaldet. Dementsprechend gab es nur passende Standorte für Bergbartfaden und Roten Hornisten. Zu jener Zeit mag Bastardierung zwischen den beiden Arten gelegentlich vorgekommen sein, war aber von geringer Bedeutung, weil kein Raum vorhanden war, an dem sich Populationen mit hybridem Ursprung hätten ansiedeln können. Mit trockener werdendem Klima wurden die Wälder durch Gestrüpp ersetzt und so neuartige Biotope verfügbar.
Einige hybride Bartfaden wanderten an einer oder mehreren Stellen in diese Lebensräume ein, und Wespen begannen sie zu besuchen. In der Folge bewirkte die natürliche Auslese eine Stabilisierung der Genkombinationen, die die vegetativen Pflanzenteile an das Leben auf den trockenen, gestrüppbedeckten Hängen und ihre Blüten an die Bestäubung durch Wespen - die häufigsten Besucher im neuen Lebensraum - anpaßten. Ein ähnlicher Verlauf der Vorgänge mag einen großen Teil der verschiedenen Bartfadenarten in den Weststaaten der USA hervorgebracht haben. Künstliche Bastardierung zwischen vielen Paaren verwandter Arten haben gewöhnlich fertile Nachkommen ergeben, so daß die gegenwärtigen Barrieren, die sie trennen, vor allem ökologische oder ökogeographische und zusätzlich Isolation durch Blütenbau sind.
Abb.15: Vier Arten
der Gattung Penstemon aus Kalifornien mit ihren
Bestäubern. Die Eindeutigkeit der Verbreitungsgebiete zeigt, dass die
extremen Typen die beiden ältesten Arten sind, und zwar Bergbartfaden
(P. Grinelli) (A) und Roter Hornist (P. centranthifolius) (B). Die beiden anderen entstanden, wie die Pfeile andeuten, durch Bastardierung und darauffolgende Isolation und Stabilisierung der intermediären Populationen durch die selektive Wirkung spezifischer Bestäuber.
Aus Stebbins 1980.
Der Hinweis, dass "an einigen wenigen Stellen" natürliche Hybride zwischen Bergbartfaden und Rotem Hornisten gefunden wurden, zeigt bereits, dass die Isolation durch die Bestäuber keine vollständige ist. Grant berichtet über diese beiden Arten und P. spectabilis 1981, p. 132:
The species are interfertile in the experimental garden. In the wild they hybridize in some areas of sympatric contact but remain isolated in others.
Was wir hier festhalten möchten, ist die Tatsache, dass es sich auch bei diesem Beispiel genetisch betrachtet um Populationen von Rekombinanten einer einzigen Art handelt: Die Ökotypen sind miteinander kreuzbar und bringen fertile Nachkommen hervor. Grundsätzlich gehören alle miteinander kreuzbaren, fruchtbare Nachkommen hervorbringende Rekombinanten zur selben Art (vgl. oben die Ausführungen zum Rekombinationsquadrat in Verbindung mit der ökologischen Isolation p. 80). Wie das Beispiel des Bartfadens zeigt, garantieren selbst ökologische, ökogeographische und zusätzliche Isolation im Blütenbau noch keine genetische Barriere. Dasselbe trifft auf die oben (p. 88) von Heß zitierten Beispiele der Ophrys-Arten zu. Gerade die Orchideen zeichnen sich durch eine erstaunliche Kreuzbarkeit nicht nur der (systematischen) Arten, sondern auch der Gattungen aus (vgl. p. 304).
Sehen wir uns das Beispiel von Ophrys näher an. Heß bemerkt, dass die einzelnen Ophrydeen-Arten primär durch ihren artspezifisch verschiedenen Blütenduft voneinander ethologisch isoliert sind. Erst später kommen dann morphologische und mechanische Kriterien ins Spiel. Baumann und Künkele stellen in ihrer ausgezeichneten Arbeit über DIE WILDWACHSENDEN ORCHIDEEN EUROPAS zur Hybridisierung folgendes fest (1982, p. 410):
Die scheinbar perfekte Anpassung der Ophrys-Blüte an die spezifischen Bestäuber kann jedoch nicht verhindern, daß die Hybridisierung bei dieser Gattung weit verbreitet ist. Normalerweise handelt es sich um Einzelbastarde. Nur selten kommen sie in größerer Zahl vor. In wenigen Fällen gibt es sogar völlig stabilisierte Hybridpopulationen, die einen eigenständigen Eindruck erwecken und ein relativ großes Areal besiedeln, wobei eine oder sogar beide Elternarten völlig verschwunden sein können. Ein schönes Beispiel dafür stellt O. delphinensis dar, eine Sippe, die durch Bastardierung zwischen O. argolica und oestrifera hervorgegangen ist. Weit verbreitet ist dies bei O. bertolonii, bei der sich eine ganze Reihe von Bastardpopulationen gebildet hat und die infolge einer weiträumigen Trennung auch als eigenständige Arten aufgefaßt werden (O. benacensis, bertoliniformes, catalaunica).
Um dem Leser einen Eindruck zu vermitteln, was sich bei der Gattung Ophrys an Hybridisation abspielt, habe ich nur einmal die Hybriden bzw. Hybridpopulationen der Arten mit den ersten drei Buchstaben des Alphabets nach Baumann und Künkele aufgeführt. Die vollständige Liste würde mehrere Seiten umfassen.
OPHRYS-BASTARDE
|
Ophrys aesculapii |
x |
O. argolica |
|
|
x |
O. ferrum-equinum |
|
|
x |
O. sphegodes |
|
Ophrys apifera |
x |
O. bertolonii |
|
|
x |
O. candica |
|
|
x |
O. holosericea |
|
|
x |
O. insectifera |
|
|
x |
O. oestrifera |
|
|
x |
O. scolopax |
|
Ophrys arachnitiformes |
x |
O. araneola |
|
|
x |
O. atrata |
|
|
x |
O. bertolonii |
|
|
x |
O. bombyliflora |
|
|
x |
O. fusca |
|
|
x |
O. holosericea |
|
|
x |
O. lunulata |
|
|
x |
O. scolopax scolopax |
|
|
x |
O. tenthredinifera |
|
|
x |
O. sphegodes |
|
|
x |
O. vernixia |
|
Ophrys araenola |
x |
O. arachnitiformes |
|
|
x |
O. atrata |
|
|
x |
O. bertolonii |
|
|
x |
O. holosericea |
|
|
x |
O. insectifera |
|
|
x |
O. sphegodes |
|
Ophrys argolica |
x |
O. aesculapii |
|
|
x |
O. mammosa |
|
|
x |
O. oestrifera |
|
|
x |
O. spruneri |
|
|
x |
O. tenthredinifera |
|
|
x |
O. vernixia |
|
Ophrys altantica |
x |
O. dyris |
|
|
x |
O. fusca |
|
|
x |
O. lutea |
|
Ophrys atrata |
x |
O. araenola |
|
|
x |
O. arachnitiformes |
|
|
x |
O. bertolonii |
|
|
x |
O. bombyliflora |
|
|
x |
O. garganica |
|
|
x |
O. exaltata |
|
|
x |
O. fusca |
|
|
x |
O. lunulata |
|
|
x |
O. oestrifera oestrifera |
|
|
x |
O. scolopax scolopax |
|
|
x |
O. sphegoides |
|
|
x |
O. tenthredinifera |
|
|
x |
O. vernixia |
|
Ophrys attica |
x |
O. bombyliflora |
|
|
x |
O. oestrifera oestrifera |
|
|
x |
O. reinholdii |
|
Ophrys bertolonii |
x |
O. apifera |
|
|
x |
O. araneola |
|
|
x |
O. arachnitiformes |
|
|
x |
O. bombyliflora |
|
|
x |
O. fusca |
|
|
x |
O. holosericea |
|
|
x |
O. insectifera |
|
|
x |
O. lunulata |
|
|
x |
O. lutea |
|
|
x |
O. scolopax |
|
|
x |
O. sphegodes |
|
|
x |
O. tenthredinifera |
|
|
x |
O. vernixia |
|
Ophrys bombyliflora |
x |
O. arachnitiformes |
|
|
x |
O. atrata |
|
|
x |
O. attica |
|
|
x |
O. bertolonii |
|
|
x |
O. doerfleri |
|
|
x |
O. ferrum-equinum |
|
|
x |
O. holosericea |
|
|
x |
O. lutea |
|
|
x |
O. oxyrrhynchos |
|
|
x |
O. scolopax scolopax |
|
|
x |
O. sphegodes |
|
|
x |
O. tenthredinifera |
|
|
x |
O. vernixia |
|
Ophrys bornmuelleri |
x |
O. holosericea |
|
|
x |
O. mammosa |
|
|
x |
O. umbilicata |
|
Ophrys caudica |
x |
O. apifera |
|
|
x |
O. holosericea |
|
|
x |
O. tenthredinifera |
|
Ophrys cilicica |
x |
O. straussii |
usw. durch das ganze Alphabet.
Von den insgesamt 51 aufgeführten Ophrys-Arten wird nur für 7 angegeben, dass bisher noch keine Bastarde nachgewiesen wurden. Sieht man sich diese 7 Arten näher an, so fällt einem zunächst auf, dass 5 von diesen Arten ein ausgesprochen kleines geographisches Areal besiedeln:
Ophrys flavomarginata: nur Zypern und Nordisrael
Ophrys hebes: nur Mittel-Griechenland
Ophrys isaura: nur kleines Verbreitungsgebiet in der südlichen Türkei (gegenüber Zypern)
Ophrys kotschyi: nur Zypern
Ophrys omegaifera: nur auf Kreta und einigen griech. Inseln
Man muss dabei berücksichtigen, dass "die europäischen Orchideen-Arten in aller Regel große Verbreitungsgebiete" besiedeln (Baumann und Künkele, p. 9). Zu Ophrys oestrifera bemerken dieselben Autoren zum Stichwort Bastarde (p.254): "Bislang keine sicheren Nachweise bekannt, da die Eigenständigkeit dieser Unterart erst jetzt erkannt wurde." Für die zuerst beschriebene Art Ophrys oestrifera oestrifera werden jedoch Bastarde mit O. apifera, argolica, attica, holosericea, sphegodes, spruneri und tenthredinifera angegeben. Ophrys schulzei schließlich besitzt ein größeres Verbreitungsgebiet als die 5 oben aufgeführten Arten, und zwar in der Südosttürkei und den angrenzenden Ländern.
Es versteht sich von selbst, dass für weitverbreitete Arten bei häufiger Arealüberschneidung mit anderen Arten sowohl die Bastardierungschancen als auch die Wahrscheinlichkeit, Bastarde in der Natur aufzufinden, größer ist als für Arten mit einem kleinen geographischen Areal. Die Behauptung, dass auch die wenigen Arten, für die bisher noch keine Bastarde beschrieben worden sind, ebenfalls mit anderen Arten bastardierbar sind, wird kein besonderes Wagnis darstellen; zumindest dürfte das für die Mehrzahl der Fälle zutreffen.
Sehen wir uns zur Hybridbildung bei Ophrys noch einige Details näher an. Ehrendorfer berichtet 1980, p. 18:
Am besten dokumentiert ist die hybridogene Sippenbildung bei Ophrys im Verwandtschaftskreis von O. bertolonii, O. atrata (O. sphegodes agg.) und ihren Zwischenformen, die man ebenso wie O. dalmatica aus praktischen Gründen dem O. bertolonii-Aggregat zurechnen kann.
Nach Hinweis, dass zwei Arbeitsgruppen unabhängig voneinander zu den gleichen Ergebnissen gekommen ist, schreibt er (p. 20):
In Süditalien sind O. bertolonii und O. atrata weit verbreitet und vielfach sympatrisch, aber durch etwas verschiedene Standortansprüche, nur wenig überlappende Blütezeit und vor allem auch durch teilweise unterschiedliche Bestäuber ihrer Sexualtäuschblumen voneinander einigermaßen isoliert. So berichten Kullenberg (1973) bzw. Kullenberg & Bergström (1974, 1976) über Blütenbesuch von Männchen verschiedener Andrena-Arten auf O. atrata, dagegen von Campsoscolia ciliata (männlich) auf O. bertolonii, während eine dritte Hymenopterengattung, nämlich Eucera nigrilabis (männlich), von beiden Ophrys-Arten angelockt wird. Diesen Beobachtungen entspricht es, daß in Süditalien im allgemeinen nur vereinzelte Bastarde, seltener individuenreiche und dann sehr variable Hybridschwärme auftreten, in denen die Kohäsion der Parentalmerkmale noch sehr deutlich ist (Abb. 4). Am Gargano ist unter starker Kanalisierung der Variabilität dem Auftreten gewisser morphologischer Sonderentwicklungen und einer Abschwächung der parentalen Merkmalskohäsion aus solchen Hybridschwärmen eine durchaus selbständige und lokal häufige Sippe entstanden, die O. & E. Danesch O. bertoliniformis genannt haben (Abb. 5).
Die Frage der Isolationsmechanismen und Hybridbildung lässt sich bei der Gattung Ophrys sowie zahlreichen weiteren Beispielen sicher noch seitenweise weiter dokumentieren. Für unsere Frage nach dem Artbegriff aufgrund präzygotischer Isolationsmechanismen lässt sich anhand der oben aufgeführten Tatsachen aber jetzt schon feststellen, dass weder Biotopisolation noch temporäre, ethologische und mechanische Isolation ausreichen, um zwangsläufig eine konstitutionell-genetische Barriere aufzubauen. Statt dessen ist Hybridbildung sogar in der Natur die Regel und Populationen von bestimmten Rekombinanten sind als eigene Arten beschrieben worden. Es ist dabei nicht uninteressant, wie die Orchideen-Forscher Baumann und Künkele den Artbegriff definiert haben. Zunächst stellen sie (1982, p. 9) fest, dass Linné die gesamte Ophrys-Fülle noch als eine einzige Art mit 8 Varietäten, nämlich Ophrys insectifera L. beschrieb und meinen dann, dass der Artbegriff sich heute nicht mehr "so eng" fassen lässt. Wenn ich die Bedeutung der Adjektive "eng" und "weit" richtig verstehe, meinen die Verfasser, dass man den Artbegriff heute nicht mehr so weit fassen sollte; denn wenn Linné die Mannigfaltigkeit der Ophrydeen-Arten (soweit damals bekannt) noch zu einer einzigen Art zusammenfasste, dann hat er auch seinen Artbegriff sehr weit gefasst; hätte er ihn enger gefasst, dann hätte er aus jeder Sippe eine eigene Art gemacht und zumindest die 8 Varietäten mit eigenen Artnamen belegt. Die Autoren schreiben weiter (p. 9):
Wir selbst gehen bei der Frage, was unter einer Art zu verstehen ist, mehr von praktischen Gesichtspunkten aus: Die Sippe muß gut im Gelände ansprechbar sein, sich also durch eine Reihe konstanter Merkmale deutlich von den nächstverwandten Sippen unterscheiden."
Hier haben wir wieder unseren morphologischen Artbegriff. Wie oben anhand der Hybriden und Hybridpopulationen gezeigt, handelt es sich genetisch sehr wahrscheinlich bei den 51 von den Verfassern in ihrem Werk aufgeführten Ophrys-Arten tatsächlich nur um eine einzige Art mit (mindestens) 51 verschiedenen Rassen, über welche uns die Autoren einen schönen Überblick vermitteln.
Oben habe ich Ehrendorfer (mit Kullenberg und Bergström) zitiert: Die Hymenoptere Eucera nigrilabis wird von den beiden Ophrys-Arten O. atrata und O. bertolonii angelockt, andere Hymenopteren jeweils nur von der einen oder der anderen Ophrys-Art. Macht man den Artbegriff bei Ophrys von der Bestäubung durch bestimmte Insektenarten abhängig, so ergibt sich folgendes Bild: Wo immer eine Insektenart von mehreren Ophrys-Arten (d.h. Rekombinanten) angelockt wird, kommt auch starke Hybridisierung vor und eine morphologische Artabgrenzung ist innerhalb einer Population oder Sippe nicht durchführbar. Folge: Verschmelzung zweier (oder mehrerer) Arten, die anderswo unterschieden werden können, zu einer Art. Verschwinden jedoch aus irgendwelchen Gründen (sagen wir besondere Empfindlichkeit gegen ein bestimmtes Pestizid) solch nichtspezialisierte Insekten aus einem Areal und verbleiben nur die bestimmte Rekombinanten anfliegenden Bestäubungs-Spezialisten in diesem Gebiet, so spaltet die eine Art in viele Arten auf. Kehren die Nicht-Spezialisten zurück, dann kommt es wieder zur Verschmelzung der Arten, d.h. wir machen schließlich Vorkommen, Häufigkeit und Verhalten bestimmter Insektenarten zum Maßstab für unseren Artbegriff bei Ophrys.
Im Übrigen treffen die meisten der oben zitierten Einwände gegen einen Artbegriff, nach dem aufgrund präzygotischer Isolationsmechanismen eigene Arten unterschieden werden (vgl. die Beispiele der Sperlinge, Eichen, Nelkenwurz, Kröten, Zebrafinken, Silber- und Heringsmöwe, Haustiere und des Menschen) auch auf die Gattung Ophrys zu.
Fazit: Populationen von unterschiedlichen, aber miteinander potenziell fruchtbaren Rekombinanten können genetisch nicht als eigene Arten geführt werden.
Oder positiv formuliert: Unabhängig von ökologischen, temporären, ethologischen und mechanischen Isolationsmechanismen, gehören alle miteinander fruchtbaren Populationen auch der unterschiedlichsten Rekombinanten zur selben Art. Dasselbe trifft auf unterschiedliche, geographisch voneinander getrennte Populationen zu.
Es wurde schon wiederholt darauf aufmerksam gemacht, dass die Überbewertung präzygotischer Isolationsmechanismen als ausreichende Arttrennungsfaktoren ihre Ursache in der neodarwinistischen Zielsetzung findet, möglichst zahlreiche Beispiele für im Werden begriffene Arten ("evolution in action") zu finden und mit dieser "Evolution" die Art und Weise des Ursprungs aller Lebensformen zu implizieren.