6) Beweisen die paläobotanische Befunde, dass der Species keine festen Grenzen gesteckt sind?
(Fortsetzung meines Briefes vom 23.8.1994.)
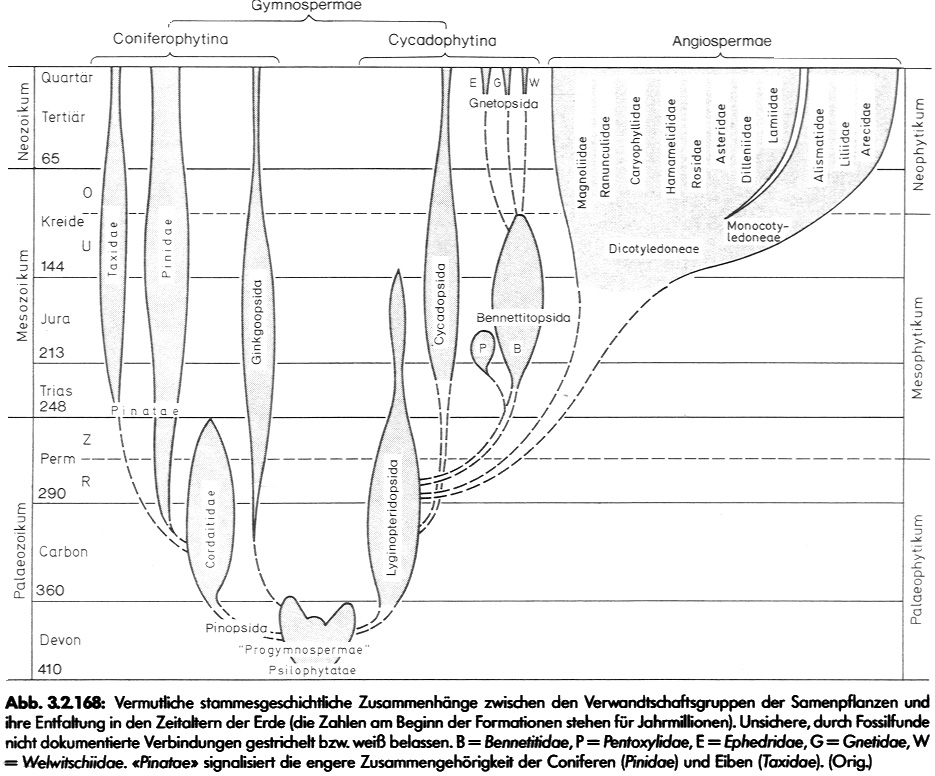
(Prof. C.:) "In der aufsteigenden Evolutionsreihe, über die ich in Köln Vogelsang referiert habe, von den frühen, den Urlandpflanzen (Rhyniophyta) noch ähnlichen Progymnospermen mit Iso- bzw. Heterosporen über die spermienbildenden und weiter siphonogamen Gymnospermen mit Samenbildung bis hin zu den hochspezialisierten Angiospermen mit ihrer gewaltigen Variationsbreite und Blütenspezialisation, kann ich nirgends "Megasaltationen" oder unüberbrückbare "Bauplanzäsuren" sehen: nur viele kleine Schritte, wie sie auch beim Vergleich verwandter Arten in Erscheinung treten."
(W.-E.L.:)
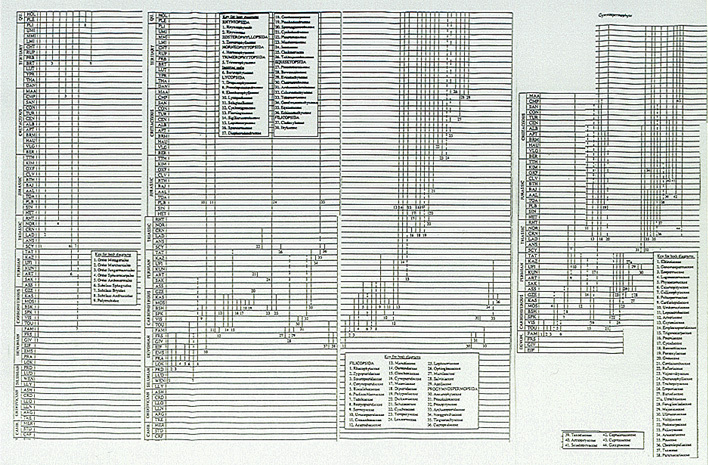
Prüfen wir anhand dieser Aussagen, ob die Behauptung von S. N. Salthe: "...metaphysical uniformitarianism is part and parcel of pure neo-Darwinism, and one of its severe weaknesses (J. Hered. 78, 213-214) korrekt ist, indem wir uns einige relevante Daten näher ansehen. (Eine erschöpfende Aufführung und Diskussion aller bislang festgestellten realen und scheinbaren Diskontinuitäten würde eine Arbeit von mindesten 500 Seiten erfordern.)Abb. 7
Fossilbericht der Moose, 'Farne' und Gymnospermen. Zusammengestellt nach Benton (1993): The Fossil Record 2. Zu Detailfragen vgl. das beigelegte DINA3 Format der Abbildung.
(N.: The original charts are much larger and very clear in contrast to this strongly miniaturized (internet-)figure; so for the details and original clarity, see the original work.)
Alle Familien und Gruppen treten sprunghaft auf; keine einzige ist durch "viele kleine Schritte", d.h. durch eine Serie von Übergangsformen mit der (morphologisch) nächsten verbunden.
Abb. 8
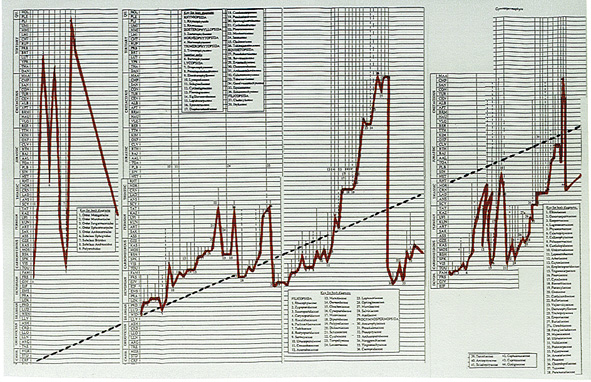
Kontrast zwischen evolutionistischer Erwartung und paläobiologischen Befunden: Wieder der Fossilbericht der Moose, 'Farne' und Gymnospermen nach Benton (1993). Kurven: Gestrichelte schwarze Linie: Ursprünglich erwartete Evolutionskurve (kontinuierliche Entwicklung). Durchgezogene rote Linie: Paläobotanische Tatsachen. Für die Details vgl. wieder die DINA3 Abbildungen.
(N.: The original charts are much larger and very clear in contrast to this strongly miniaturized (internet-)figure; so for the details and original clarity, see the original work.)
Abb. 9 (folgend):

Wie Abb. 2, nur die Kurven etwas gerundet ("ausgeglichen") und die evolutionistischen Erwartungen leicht modifiziert (gestrichelte und rote Linien). Auch bei weiteren halbwegs realistischen Modifikationen der Linien bleibt der Kontrast zwischen kontinuierlicher Evolution und den paläobiologischen Realitäten unübersehbar!
(N.: The original charts are much larger and very clear in contrast to this strongly miniaturized (internet-)figure; so for the details and original clarity, see the original work.)
Auf den vorausgehenden Abbildungen ist das jeweils erste und letzte Auftreten der verschiedenen Familien und Gruppen der Moose, 'Farne' und Gymnospermen nach dem heutigen Stand der paläobiologischen Forschung zusammengestellt. Nach Benton (1993) : The Fossil Record 2 (Chapman & Hall). In der ersten Abb. sind nur die Daten aufgeführt und in den folgenden Abbildungen der Kontrast zwischen der ursprünglich erwarteten Evolutionskurve (Hofmeistersche Serie) - zunächst auch von der Synthetischen Evolutionstheorie übernommen und später modifiziert - und den bisherigen paläobiologischen Tatsachen. (Die stark verkleinerten Abbildungen sind zur besseren Lesbarkeit im DINA3 Format noch einmal beigelegt.)
Es hat Jahrzehnte gebraucht, bis sich die Erkenntnis allgemein durchgesetzt hatte, dass eine Evolution des Pflanzenreichs im Sinne der Hofmeisterschen Serie nicht haltbar war. Da nun ein einziger Stammbaum zur Erklärung der Phänomene nicht mehr ausreichte, hat man sich heute damit beholfen, den Stammbaum in einen Stammbusch zu verwandeln: Die Rhyniophyta stammen danach nicht von Moosen, sondern sowohl die Rhyniophyta als auch die Moose von noch unbekannten Algen ab. Das Prinzip der konvergenten Entwicklung wurde dann auf die verschiedensten Pflanzengruppen ausgedehnt mit dem Resultat, dass nach der heutigen Auffassung eine ganze Anzahl hochkomplexer Strukturen immer wieder 'von neuem erfunden' wurde (vgl. die sich widersprechenden 'Stammbäume' S.27 -29 des Briefes).
Das Thema 'Konvergenz im Pflanzenreich' wäre nun wieder ein umfangreiches Kapitel für sich. Ich möchte an dieser Stelle nur bemerken, dass die Idee der unabhängigen und oftmals wiederholten Entwicklung von hochkomplexen (genetischen, physiologischen und anatomisch-morphologischen) Systemen durch das neodarwinistische Faktorensystem (definitionsgemäß richtungslose 'Kleinmutationen', Selektion und Isolation etc., d. h. durch viele kleine Veränderungsschritte in verschiedenen Bereichen mit daraus resultierenden zahllosen fließenden Übergängen) die gesamte Abstammungslehre immer unwahrscheinlicher macht. Denn war es schon unwahrscheinlich genug und erforderte es viel 'Glauben', die Entstehung einer komplexen Struktur oder eines synorganisierten Systems durch dieses Faktorensystem auch nur einmal für möglich zu halten, so wird jetzt sogar 'der Glaube' gefordert, dass komplexe Systeme mehrmals unabhängig voneinander durch 'Zufallsmutationen' und Selektion entstanden sein sollen. Eine genauere biologische und mathematische Untersuchung der aus dem Konvergenzpostulat folgenden, sich potenzierenden Unwahrscheinlichkeiten könnte einen bedeutenden Beitrag zur Entmythologisierung der Evolutionstheorie liefern.
Kennzeichnend für die paläobiologischen Befunde ist das durchweg sprunghafte Auftreten neuer Gruppen sowie die außerordentliche (evolutionistisch überhaupt nicht erwartete) Konstanz vieler Formen oft über Hunderte von Jahrmillionen hinweg (und ebenso das abrupte Verschwinden zahlreicher Gruppen). Die - besonders bei höheren systematischen Kategorien - schwere Diskrepanz zwischen der Idee der kontinuierlichen Evolution ("in vielen kleinen Schritten" ohne "Megasaltationen" und unüberbrückbaren "Bauplanzäsuren") und den paläobiologischen Befunden kennzeichnet fast die gesamte Geschichte der Paläontologie von 1859 bis auf den heutigen Tag (vgl. Gould und Eldredge 1993: Punctuated equilibrium comes of age. Nature 366, 223-227, dort umfangreiche weitere Literaturangaben). Der Paläontologe Kuhn fasste diese Diskrepanz einmal wie folgt zusammen:
"Das Vorurteil, dass die Stammesgeschichte nur eine Summierung kleinster Abänderungsschritte sein könne und bei entsprechend vollständiger Kenntnis der paläontologischen Urkunden die kontinuierliche Entwicklung zu beweisen sei, ist sehr tief eingewurzelt und weit verbreitet. Aber die paläontologischen Tatsachen sprechen schon lange gegen dieses Vorurteil! Gerade deutsche Paläontologen wie Beurlen, Dacque und Schindewolf haben mit Nachdruck darauf hingewiesen, dass aus vielen Tiergruppen ein so reiches, ja geradezu erdrückendes fossiles Material vorliegt (Foraminiferen, Korallen, Brachiopoden, Moostiere, Cephalopoden, Ostracoden, Trilobiten usw.), dass man die nach wie vor zwischen den Typen und Subtypen bestehenden Lücken als primär vorhanden auffassen muss"
(Kursiv vom Verfasser).Vgl. dazu die Dokumentation in der Artbegriffsarbeit pp. 298, 314-323, 593, 594.
Inwieweit treffen Kuhns Feststellungen auch auf die Paläobotanik zu? Heribert Nilsson, Professor für Botanik an der Universität Lund, hat 1953 (3 Jahre nach Stebbins' Variation and Evolution in Plants) dieser Frage ein umfangreiches Kapitel gewidmet (Synthetische Artbildung, pp. 419 -506), in dem er zu folgendem Schluss kommt (Zusammenfassung S. 1212):
"It may, therefore, be firmly maintained that it is not even possible to make a caricature of an evolution out of the palaeobiological facts. The fossil material is now so complete that it has been possible to construct new classes and the lack of transitional series cannot be explained as due to the scarcity of the material. The deficiencies are real, they will never be filled."
Hat sich die Situation inzwischen grundlegend geändert? Die Antwort habe ich mit den Paläobotanikern Thomas und Spicer schon in der Artbegriffsarbeit gegeben (vgl. S. 320). Der Einfachheit halber sei ihre Antwort hier wiederholt. Sie stellen 1987, p. 65, in ihrer Arbeit The Evolution and Paleobiology of Land Plants fest:
"If evolution was a gradual process, one would expect to find that the fossil record would tell a story of gradual change of one form into another. Everywhere there would be intermediate forms connecting one 'species' with another. Unfortunately, the fossil record is characterized by an overwhelming absence of such intermediates. 'Missing links' are the rule rather than the exception.
Darwin considered the fossil record as being particularly important and yet it singularly failed to support his gradualist ideas. The gaps between species were glaringly wide and despite continued research they remain so to this day.
...over a century of investigations has failed in most cases to narrow significantly those tantalizing gaps. The nature of the fossil record is essentially one of suites of specimens that are morphologically distinct from one another. The record is punctuated by discontinuities.
The evolutionary branches are not attached to the evolutionary tree" (kursiv von mir).
Desgleichen zeigt ein gründliches Studium der Ausführungen Nilssons mit daran anschließendem Vergleich der Abbildung 1 - 3 dieses Briefes und weiteren paläobiologischen Daten, dass zwar einige (bis auf die Progymnospermen meist recht geringfügige) Korrekturen notwendig sind, dass die generelle Diskrepanz zwischen der Idee der kontinuierlichen Evolution und den paläobotanischen Befunden jedoch nicht nur nicht beseitigt ist, sondern an vielen Stellen durch weitere Funde noch zugenommen hat. So ist Nilssons Argumentation gegen die Moose als Bindeglieder zwischen Algen und Farnen wie überhaupt gegen die Hofmeistersche Serie als Abstammungsfolge inzwischen weitgehend akzeptiert (s. o.) und sowohl zwischen Algen und Rhyniophyta einerseits und Algen und Moosen andererseits besteht - das sei besonders betont - eine immense Kluft, die man sehr gut begründet in die Kategorie der "Megasaltationen" und/oder Bauplanzäsuren einordnen kann. Das nach der Theorie der kleinen Schritte völlig unerwartet frühe Auftreten der Bärlappgewächse mit Baragwanathia longifolia wird inzwischen ebenfalls von den meisten Autoren - wenn auch nach jahrelangen Auseinandersetzungen - akzeptiert.
Wie schon erwähnt, repräsentieren die Moose selbst auch eine Gruppe, die wir nach Goebel 'weder nach oben noch nach unter an andere anknüpfen können' (Möbius: Geschichte der Botanik). "Beide Klassen der Moose, die Lebermoose (Hepaticae) wie die Laubmoose (Musci), sind wesentlich höher organisiert als die höchststehenden Algen…Wir kennen bis jetzt weder lebende noch fossile Übergangsformen" (Mägdefrau 1959, p. 309; Heberers Evolution der Organismen). "Historically, the hornworts and liverworts have been interpreted as a group phylogenetically intermediate between the green algae and vascular plants (Smith 1938). Others have suggested that in general the bryophytes represent examples of evolutionary failures, or perhaps originated from early vascular plants like the rhyniophytes through loss of vascular tissue. Today, however, these views are changing based on a variety of ultra structural, biochemical, and molecular data that suggest that the principal bryophyte groups had separate origins (Crandall-Stother, 1980)" (Taylor and Taylor 1993, pp. 147/148; The Biology and Evolution of Fossil Plants; kursiv von mir).
Hier erhebt sich die Frage, wo diese separaten Ursprünge durch die behaupteten 'vielen kleinen Schritte, wie sie auch beim Vergleich verwandter Arten in Erscheinung treten', zu finden sind. A.J.E. Smith bemerkt in seiner Monographie The Liverworts of Britain and Ireland (1991, S. 6):
"Nothing is known of the phylogeny of liverworts or hornworts, although this has been the subject of much speculation, and the following classification should be regarded as natural (or phenetic) and no phylogenetic conclusions should be constructed from it."
Abgesehen von vielen Vermutungen zu den Laubmoosen (z.B. Bresinsky 1991; Strasburger) gibt es bislang keine realen Zwischenformen, die die Bauplanunterschiede zwischen Moosen und Algen durch viele kleine Schritte überbrücken würden, d .h. biologisch real nachgewiesen sind bisher nur die Bauplanzäsuren, nicht aber die postulierten vielen kleinen evolutionistischen Schritte durch 'Mikromutationen' mit "slight or even invisible effects on the phenotype". Das Kontinuum ist das 'metaphysische' Postulat (um Salthes Begriff zu gebrauchen), die immensen Bauplanunterschiede die biologische Realität.
Wir haben damit bisher schon drei ausgezeichnete Kandidaten für unüberbrückbare Bauplanzäsuren erwähnt: 1) Die Entstehung der 'Urlandpflanzen', und 2) und 3) die unabhängige Entstehung der Leber- und der Laubmoose. (Nach M. Ingrouille (Diversity and Evolution of Land Plants 1992, S. 85) sogar vier: Liverworts, hornworts and mosses erscheinen in seinem Stammbaum unverbunden.)
In keinem dieser grundlegenden Beispiele kann man 'nur viele kleine Schritte (als reale Zwischenformen) sehen, wie sie auch beim Vergleich verwandter Arten in Erscheinung treten'. Eine kontinuierliche Entwicklung im Sinne des Neodarwinismus kann man zwar postulieren und auch fest daran "glauben". Zu sehen ist hingegen die Tatsache, dass die Bindeglieder seit über 100 Jahren fehlen und dass viele dieser erwarteten Übergangsformen sowohl selektionstheoretisch als auch genetisch, physiologisch und morphologisch-anatomisch derart unwahrscheinlich sind, dass man das Postulat ihrer einstigen Existenz auch mit guten wissenschaftlichen Gründen bezweifeln kann.Inwieweit trifft das nun auch auf die von Herrn Prof. C. zitierten "Progymnospermen mit Iso- bzw. Heterosporen über die spermienbildenden und weiter siphonogamen Gymnospermen mit Samenbildung bis hin zu den hochspezialisierten Angiospermen" zu?
Zunächst zu den "Progymnospermen" (eine nicht ganz glückliche Wortprägung, da sie die Theorie z. T. schon vorwegnimmt): Der wichtigste und grundlegendste Unterschied zwischen den Trimerophytopsida und Progymnospermen (wie Aneurophyton und Rellimia) "is the presence of secondary vascular tissues produced by a bifacial cambium in the progymnosperms" (Steward/Rothwell 1993, p. 275; Paleobotany and the Evolution of Plants). "The lignophyte clade is defined by the presence of a vascular cambium that produces secondary xylem centripetally and secondary phloem centrifugally" (Crane 1989, S. 166; Allen/Briggs; eds.: Evolution and the Fossil Record).
Damit haben wir den nächsten Kandidaten für eine durch Mikroevolution vermutlich unüberbrückbare "Bauplanzäsur". Eine für den Bau und die Entwicklungsmöglichkeiten der Pflanze durchgreifende genetische, anatomische und physiologische "Innovation" (Crane) ist festzustellen: das bifaziale, sekundäres Xylem und sekundäres Phloem bildende Cambium. Diese geniale Erfindung und erdgeschichtlich revolutionäre Neuheit setzt sehr wahrscheinlich eine ganze Batterie von neuen in Sequenz und Funktion genauestens aufeinander abgestimmten Struktur- und Regulatorgenen voraus (zur Wahrscheinlichkeit der Entstehung spezifischer DNA-Sequenzen durch die definitionsgemäß richtungslosen Zufallsmutationen vgl. wieder Artbegriff S. 359). Genau an diesem Punkt könnte man (ganz vorsichtig formuliert) vielleicht auch einmal den Gedanken in Erwägung ziehen, 'dass Arten mit Gewinn an genetischer Information, mit Neukonstruktionen samt Synorganisationen auf den verschiedensten Ebenen durch einen intelligenten Schöpfungsakt entstehen können' (vgl. das Zitat Artbegriff S. 591, dort auch Seitenangaben zu weiteren Ausführungen).
Auch die realgenetische Ableitung der verschiedenen Grade der Heterosporie von isosporen Formen könnte sich als komplizierter erweisen als man sich das im allgemeinen heutzutage vorstellt. Zudem müsste die Heterosporie vielfach unabhängig voneinander entstanden sein. Ich kenne bislang kein einziges Beispiel, dass bei isosporen Formen durch Mutationen Heterosporie induziert worden wäre. Es wäre jedenfalls sicher aufschlussreich, die hinter dieser Fragestellung stehenden genetischen und molekularen Ursachen miteinbeziehen zu können.
Wie sieht es nun mit der weiteren Entwicklung zu den Gymnospermen und Angiospermen aus? Finden wir hier ein Kontinuum über viele kleine Schritte? Wenn dem so wäre, dann müssten heute alle wesentlichen Abstammungsfragen bis zur Herkunft der Angiospermen gelöst sein, und alle Biologen würden diese Fragen aufgrund der fossilen Befunde in völliger Übereinstimmung beantworten können.
Mehr brauche ich Biologen, die sich mit den gegenwärtigen Kontroversen dieser Thematik auskennen, eigentlich gar nicht zu sagen. Aber gehen wir noch etwas ins Detail. Crane bemerkt (1989, S. 167): "While it is generally accepted that the progymnosperm grade includes those pteridophytes most closely related to seed plants, relationships between aneurophytes, archaeophytes and seed plants are controversial" (kursiv von mir); und er fährt fort:
"Initially the demonstration of heterospory in some archaeopterids indicated that they may be the closest group to spermatophytes (Pettitt 1970; Meyen 1984), but more recently the recognition of anatomical similarities between the earliest seed plants and aneurophytes has supported the opposite conclusion (Rothwell and Erwin 1987). Beck (198l) has suggested a biphyletic origin of seed plants with cordaites and conifers derived from archaeopterids, and Palaeozoic 'seed ferns' (see below) being derived from aneurophytes."
Ähnlich bemerken Taylor und Taylor über die Progymnospermen 1993, S. 459; Lit. s.o.):
"Since the initial establishment of the group, there has been widespread agreement that the progymnosperms are the ancestral plexus from which the gymnosperms evolved. However, opinions vary as to whether the gymnosperms all evolved from a common ancestor or whether the various gymnosperm groups can be traced to more than one ancestor. In a very real sense, the issue revolves around whether the seed habit evolved only once or had multiple origins.
One hypothesis suggests that the Aneurophytales are the ancestral progymnosperm group from which the gymnosperms evolved. In particular, it has been suggested that the aneurophytes can be traced to the Lower Carboniferous seed ferns, and from one or more of these groups, the remaining gymnosperms evolved (Rothwell, 1982a). Support for this hypothesis includes the discovery of Upper Devonian cupulate seeds together with vegetative axes that possess the frond architecture of Lower Carboniferous pteridosperms (Rothwell and Erwin, 1987). Anatomically, however, the vascular system of these axes is protostelic like that of aneurophytes, suggesting that these Upper Devonian plants are transitional between progymnosperms and early seed ferns.
A different view of the origin of gymnosperms is presented by Beck in a series of papers summarized by Beck and Wight (1988). According to this interpretation, gymnosperms are polyphyletic, with the seed ferns evolving from an aneurophyte ancestor, while the cordaites and conifers had their origin within the Archaeopteridales. One additional scenario that deserves mention is Meyen's (1984) suggestion that the gymnosperms are derived from the archaeopteridalean progymnosperms.
Despite the ideas presented above, we are a long way from determining the ancestral relationships of the major groups of gymnosperms. In part this reflects the paucity of information during the Upper Devonian and Lower Mississippian transition and also reflects the difficulty of interpreting certain fossils. Several workers have attempted to provide greater unbiased resolution to the question of the origin of gymnosperms using cladistic methodology (Crane, 1985a; Doyle and Donoghue, 1986). Although these analyses perhaps provide more support for a monophyletic origin for the gymnosperms, evolving the simple leaves and reproductive organs of cordaites and conifers from the compound fronds and cupulate seeds of seed ferns continues to remain a major obstacle."
Charles B. Beck und David C. Wight sind auf die Details dieser Kontroverse zusammenfassend wie folgt zu sprechen gekommen (1988, S. 68/69; Origin and Evolution of Gymnosperms, editor: Charles W. Beck; Columbia University Press):
"Despite the acceptance of a general ancestor-descendent relationship between progymnosperms and seed plants, determination of the specific ancestral taxon or taxa from which the major groups of gymnosperms evolved has remained the subject of some controversy. In a series of papers, Beck (1957,1966,1970,1971, 1981) has developed the idea that pteridosperms and their descendants evolved from some aneurophyte-like progymnosperm, whereas the cordaites, conifers, and related taxa had their origin in the Archaeopteridales. This hypothesis suggests that seed plants are a polyphyletic group - i.e., that the seed habit evolved twice. In contrast, Rothwell (1981,1982) has suggested that seed plants are monophyletic. He believes that aneurophytes represent an ancestral group from which pteridosperms evolved (a similarity with Beck's hypothesis); that "coniferopsids" are derived from within the seed fern complex, presumably from a taxon similar to Callistophyton; and that Archaeopteris, therefore, is unrelated to any group of seed plants. Meyen (1984, 1986) has presented a third alternative, in which seed plants are polyphyletic, as in the Beck hypothesis, but he considers both initial groups of seed plants to be derived from archaeopteridaleans.
Aneurophytales are attractive candidates for ancestor of the earliest seed plants. Many of the early pteridosperms - e.g., Stenomyelon, Tristichia, Tetrastichia, Microspennopteris, Heterangium, Galtiera, etc. are characterized by protosteles and secondary xylem similar to those of aneurophytaleans. In addition, similarities between aneurophytalean fertile organs and the pollen organs (Stidd and Hall 1970; Rothwell 1981) and cupulate systems of early lyginopterids (Doyle and Donoghue 1986a) have been cited as evidence of relationship. Although Meyen (1984) argues for an archaeopteridalean ancestry of seed ferns, we regard this as a somewhat less likely alternative and tentatively interpret similarities between these two groups (e.g., relatively simply secondary phloem in both and similarities between the leaf-Iike lateral branch systems of Archaeopteris and fronds of pteridosperms) to have resulted from parallel evolution.
Understanding the relationships of coniferopsids, however, has proven to be a more difficult problem. As evidence in support of an archaeopteridalean ancestry for coniferopsids, Beck (1966,1970,1971,1981) has emphasized similarities in habit, lateral branch systems, and secondary xylem between Archaeopteris and early conifers. In contrast, Rothwell (1981, 1982) has primarily emphasized similarities in reproductive biology between coniferopsids and the pteridosperm Callistophyton in support of his hypothesis. With respect to the Beck hypothesis, Rothwell notes that similarities between Archaeopteris and coniferopsids are solely vegetative, and that known features of reproductive biology are "extremely different." He also cites the stratigraphic hiatus between the most recent occurrences of Archaeopteris (Tournaisian) and the earliest occurrence of a cordaite (Namurian) or a conifer (Westphalian), and the difference in divergence of traces from cauline stelar bundles between Archaeopteris (radial) and early seed plants (tangential) as evidence against their relationship (Rothwell 1976, 1982). Meyen (1984) agrees with Beck in suggesting Archaeopteris as the ancestor of coniferopsids and the polyphyletic status of gymnosperms."
N.: Charles B. Beck und David C. Wright beschreiben die Problematik weiter wie folgt (pp. 69/70):
Several recent workers attempting to resolve the question of seed plant mono- or polyphyly and the position of aneurophytes and/or archaeopterids as ancestors have assembled data within the context of an analysis based on cladistic principles (e.g., Crane 1985a, 1985b; Doyle and Donoghue 1986a, 1986b, 1987; Stein 1986a, 1986b, 1987). These analyses are noteworthy in several respects, but primarily for the explicit framework within which characters are analyzed and taxa assigned to states. We shall consider the three hypotheses presented above (Beck, Rothwell, and Meyen) primarily within the context of the analysis of Doyle and Donoghue (1986a) because it is the most recent and, in many respects, the most explicit and complete, and because it provides a useful framework for discussion.
Doyle and Donoghue performed a strict parsimony analysis that produced a total of thirty-six equally parsimonious cladograms, or trees (their term). Most of the variation among the thirty-six trees was the result of what they considered relatively minor rearrangements of a few taxa (Doyle and Donoghue 1986a: 352-360) and pointed to the need for more morphological data on Paleozoic seed ferns. Of the thirty-six, Doyle and Donoghue chose one tree (1986a: figure 4) as their preferred phylogenetic hypothesis because, for various reasons, they consider it more likely than the others. One interesting feature of their results is that their most parsimonious cladograms support the status of gymnosperms as a monophyletic group and show coniferopsids nested somewhere within the seed fern complex, thus providing support for Rothwell's hypothesis. According to their cladograms and the character transitions shown, seed plants are descendants of a heterosporous plant that was otherwise no different from an aneurophytalean progymnosperm.
Although we believe that these results are significant, we feel, as do Doyle and Donoghue (1986a), that an archaeopteridalean origin for coniferopsids remains a viable alternative. As part of their analysis, they investigated the effects of forcing particular groups together in order to evaluate competing hypotheses of relationship among progymnosperms and seed plants. Although tree topologies consistent with Meyen's hypotheses for the origin of seed plants were considerably less parsimonious (150 steps), a cladogram consistent with Beck's hypothesis was only very slightly less parsimonious (124 steps) than their most parsimonious trees (123 steps). Given the amount of homoplasy apparently present in the data (124 character state transitions in a cladogram based on a data matrix containing 62 characters implies that, on average, each character undergoes two transitions: either two parallel evolutionary events or a single evolutionary event followed by a reversal), we consider the difference of one character state transition between the two hypotheses insignificant and an insufficient basis upon which to choose the more viable alternative. This view is reinforced by several other considerations discussed below. Some of these were also discussed by Doyle and Donoghue (1986a).
N.: Unter den weiteren Untertiteln "Lack of Information on Critical Taxa, "Missing Information on Taxa Included in the Analysis", "Taxa Excluded from the Analysis" ,"X-Coding of Characters"; "Effect of New Information on Character Coding" und "Further Consideration of the Beck and Rothwell Hypotheses" diskutieren die Autoren auf den Seiten 70-75 die verschiedenen Auffassungen bis ins Detail (diese Diskussion hatte ich ebenfalls in diesem Briefwechsel wiedergegeben; der daran interessierte Leser sei auf die Originalarbeit verwiesen).
N.: Einen wichtigen Ansatz zur Revision eines bekannten Lehrbuchbeispiels erwähnen die Autoren Charles B. Beck und David C. Wright auf der Seite 73:
"Similarly, recent work on the primary vascular systems of early seed plants and progymnosperms (Beck and Stein 1985, 1987; Wight 1985, 1986, 1987; Stein and Beck in press) has shown that protoxylem architecture can be a useful morphological feature to consider in interpreting homologies in stelar morphology. This work, along with recognition of the diversity of morphologies present among early Carboniferous "protostelic" seed plants, demonstrates that the simple transition from protostele to eustele, proposed in most phylogenetic analyses treating the origin of seed plants, may be inaccurate" (Schriftbild in diesem Satz von mir).
N.: Obwohl zum Teil nur noch von historischem Interesse, so wäre doch die gesamte Detail-Diskussion der verschiedenen Abstammungshypothesen überflüssig gewesen, wenn wir tatsächlich kontinuierliche Übergangsserien hätten! - Das ist jedoch bis heute nicht der Fall. Neuere Revisionen werden derzeit zumeist nicht auf der Basis neuentdeckter Fossilserien, sondern vor allem aufgrund molekularer Daten durchgeführt. Um ein sehr bedeutendes neueres Beispiel zu erwähnen: Die lange (auch von Doyle und Donogue mit cladistisch interpretierten morphologisch-anatomischen Merkmalen) behauptete systematische Stellung der Gnetophyten als Bindeglieder zwischen Gymnospermen und Angiospermen wurde inzwischen fast auschließlich aufgrund molekularer Vergleiche stark revidiert (G.Theißen und Mitarbeiter).
Aufgrund molekularer Daten (jedoch nicht durch den Nachweis kontinuierlicher Übergangsserien!) ist auch die Polyphylie-Hypothese für die lebenden Gymnospermen inzwischen fast ganz aufgegeben worden.
Zur Problematik der Stammbäume vgl. oben:
mendel13.htm(
Zur Ergänzung dazu sei jedoch festgestellt: auch wenn "der große Stammbaum" gar nicht existiert, so können die molekularen Daten dennoch wesentlich dazu beitragen, die konstitutionelle Verwandtschaft der Lebensformen zu klären. In diesem Sinne ist die Vielzahl der neueren (hier sogar zu einem großen Teil übereinstimmenden!) molekularen Daten zur Stellung der Gnetophyten ein wichtiger Beitrag zur Systematik, selbst wenn auch hier wieder gewisse Widersprüche zwischen Morphologie und molekularen Befunden bestehen bleiben. Vgl. Donoghue 2000.)Neuere evolutionstheoretische Literatur:
Bowe, L. M., Coat, G. dePamphilis, C.W. (2000): Phylogeny of seed plants based on all three genomic compartments: Extant gymnosperms are monophyletic and Gnetales' closest relatives are conifers. PNAS 97: 4092-4097.
Chaw, S.-M., Parkinson, C.L., Cheng, Y. Vincent, T.M. Palmer, J.D. (2000): Seed plant phylogeny inferred from all three plant genomes: Monophyly of extant gymnosperms and origin of Gnetales from conifers. PNAS 97, 4086-4091.
Donogue, M.J. (2000): Seed plant phylogeny: Demise of the anthophyte hypothesis? Current Biology 10: R106-R109.
Kuzoff, R.K., Gasser, C.S. (2000): Recent progress in reconstructing angiosperm Phylogeny. Trends in Plants Science 5: 330-336.
Qiu, Y-L., Lee,J., Bernasconi-Quadroni, Soltis, D.E., Soltis, P.S., Zanis, M., Zimmer, E.A., Chen, Z., Savolainen, V., Chase, M.W. (1999): The earliest angisoperms: evidence from mitochondrial, plastid and nuclear genomes. Nature 402: 404-407.
Winter, K.-U., Becker, A., Münster, T., Kim, J.T., Saedler, H., Theißen, G. (1999): MADS-box-genes reveal that gnetophytes are more closely related to conifers than to flowering plants. PNAS 96: 7342-7347.
Zur Problematik der Homologiefrage vgl. weiter die ausgezeichnete Diskussion von Jonathan Wells 2000, pp. 45-80 und 177-192; sowie aus der mehr neodarwinistischen Perspektive: Gregory R. Bock and Gail Cardew (1999): Homology. Novartis Foundation Symposium 222. John Wiley & Sons. Chichesterpp. 1-256 (Summary pp.243-245 by Brian Hall. - Buchrückentext u.a.: "The use of information about conserved molecules in seemingly conserved developmental processes has also proven controversial").
Wells, J. 2000): Icons of Evolution - Science or Myth? (Why much of what we teach about evolution is wrong). Regnery Publishing, Inc. Washington.
Zum Thema Monophylie und/oder Polyphylie zeichnet sich übrigens ein genereller Trend ab: Es werden (in erster Linie wieder) aufgrund molekularer Daten zu Organen und Organismen, die so unterschiedlich sind wie die verschiedenen Augentypen und die Gymnospermen, zunehmend Polyphylie-Hypothesen fallen gelassen
(auf der anderen Seite 'entstehen' dadurch jedoch auch wieder neue 'Konvergenzen' - kurz: was zum Beispiel bei den Gnetophyta als angiospermenähnlich betrachtet wurde, gilt jetzt als konvergent entstanden). Ich verstehe den Trend zur Monophylie im Rahmen des universellen Zusammenhangs aller Lebensformen im schon oben nach ReMine zitiertem Sinne:"Life is the product of a single designer - life was intentionally designed to resist all other interpretations of origin."
"This system of living objects was constructed by a single source (e.g., a common designer)."
"Look at the unifying message. It says life was designed as a unified body of objects. Not one single species is an outlander. All organisms are tied together by underlying similarity. A complex web of factors links all life. These similarities extend down to bio molecules. Biologic universals as DNA/RNA, proteins, amino acids, and ATP join life together at the molecular level."
"Similarity and diversity both have a role in the biotic message. Similarity makes life look like the work of a single designer, while diversity makes life difficult to explain by naturalistic purposes. Similarity and diversity are the alphabet of the biotic message."
"Message theory says life was designed as a biotic message. Life was designed to look like the product of a single designer (the unifying message). Yet life was also designed to resist evolutionary interpretation."
"Life's designer intentionally created life to look like a message from a single designer."
"Besides the nested pattern, the unity of life is augmented with various features which subtly knit life together more visibly than biochemicals can. For example, mimicry, mutualism, symbiotic relationships, and convergences unite the system of life into a unified whole. Darwin was able to see this unity even though he had no knowledge of the universalities at the biochemical level. This is one unified system of life; attributable to only one designer."
"The embryological similarities serve to directly unify life on a broader scale than can be observed by adult organisms alone. The system of life was designed to be unified, and the biomessage sender has gone to great lengths to ensure it is seen that way."
Auf der anderen Seite widersprechen die irreducibly complex systems und paläontologischen Diskontinuitäten etc. der Synthetischen (und anderen) Evolutionstheorie(n). - Die Aufgabe einer der gesamte biologischen Realität angemessenen Theorie liegt darin, sowohl die Ähnlichkeiten als auch die Unterschiede in einer Synthese vereinen! Die "Kreationisten" haben jedoch für ihre Theorie bisher hauptsächlich auf die Unterschiede konzentriert, die Evolutionisten hingegen auf die (homologen) Ähnlichkeiten. (N.: Ende.)
Wie schon gesagt, würde die Aufgabe, sämtliche Diskontinuitäten - von den Rhyniophyta bis zu den Angiospermen - detailliert herauszuarbeiten, eine wenigsten 500 Seiten umfassende Studie für sich beanspruchen. Aber dazu fehlt mir die Zeit. Kurz erwähnt sei noch, dass die Interpretation von der "Verwachsung von Hülltelomen zum (ersten) Integument bei Samenanlagen von A Genomosperma kidstonii (1,5 x), B G. latens (2x), C (links) Eurystoma angulare (2,5x) (alle Unterkarbon)" (Strasburger 1991, S. 725; Abbildung 3.2.183) von den Paläobotanikern Rothwell und Scheckler (1988) nicht unbedingt geteilt wird. Taylor und Taylor fassen 1993, S. 474 und S. 477 diese Frage wie folgt zusammen:
"Rothwell (1986) has pointed out some of the pitfalls in using symmetry as a diagnostic character in classification. These include the presence of a variety of shapes within specimens assigned to the same taxon and the fact that shape may vary with other factors. such as developmental level.
"It has also been suggested that the open or closed nature of the integumentary lobes in some of the earliest seeds may simply reflect stages in the maturity of the ovules, rather than stages in integument evolution ( Rothwell and Scheckler. 1988).
"These authors suggest that in several hydrasperman types integumentary lobes were open at pollen-receptive stages and closed once pollination was effected" (N.: Hervorhebung im Schriftbild von mir).
Darüber hinaus handelt es sich nach Thomas und Spicer (1987, S. 143) bei den Samenanlagen von Genomosperma zwar um die primitivsten, aber keineswegs um die ältesten, d. h. die angenommene Evolution in vielen kleinen Schritten müsste schon vorher abgelaufen sein.
Demonstriert ein Vergleich unterkarbonischer Formen von Genomospermaarten und Eurystoma nun tatsächlich "höchst eindrucksvoll die fortschreitende Fusion von Hülltelomen (bzw. steril gewordenen Megasporangien) um einen mittelständigen Nucellus (fertiles Megasporangium), also die Bildung eines (ersten) Integuments"? (Strasburger, S. 725) Dazu einige weitere Daten:
Rothwell und Scheckler berichten zusammenfassend (1988, S. 91) von 'Samenfunden', die nach gängiger geologischer Zeitrechnung bis zu 10 Millionen Jahren älter sind als die unterkarbonischen.
"Fossils and Facies For almost ten years the ovulate cupules of Archaeospenna arnoldii Pettitt and Beck (1968: figures 10-12) stood as the only bona fide evidence for a Devonian gymnosperm. Since 1977, however, there has been a greatly expanded interest in earlier gymnosperms, so that now we recognize several new genera and have reinterpreted some previously described, but poorly understood, fossils as seeds or seed-bearing cupules. These are derived from Famennian and Tournaisian Devonian deposits of North America, western Europe and Asiatic Europe (figure 2.1). Included are numerous specimens from near Elkins, West Virginia (Gillespie et al. 1981) that we shall refer to as the "Elkins" fossils; Moresnetia zalesskyi, from several deposits in Belgium; Xenotheca devonica, from southern England; and cupulate specimens described as Hydraspenna tenuis, from southern Ireland. In addition, several other fossils may represent seeds (e.g.. Spennolithus devonicus), but their taxonomic or organographic identity remains equivocal."The most ancient ovules are the Elkins specimens and Moresnetia, which are both middle Famennian (lower Fa2c; figure 2.1). They are followed by Archaeosperma (Fa2d), then Xenotheca (Fa2d-lower Tn1b); the youngest are Irish Hydraspenna and Spennolithus, which are both Tournaisian (Tn1a-lower Tn1b)."
Den unterschiedlichen Verwachsungsgrad des Integuments deuten die Verfasser bei den ältesten Funden ontogenetisch (1988, S. 101):
Abb. 10
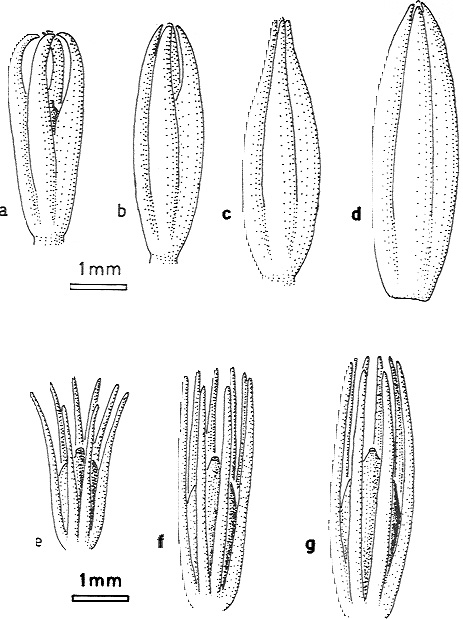
Figure 2.5. Reconstructions of ovules to show their sizes and integument morphologies at various stages inferred as pre- and post-pollination. a-d Elkins plant: a Pre-pollination to pollination configuration. b, c Post-pollination configurations. d Dispersed seed. e-g Moresnetia zalesskyi: e Pre-pollination to pollination configuration. f, g Post-pollination configurations.
(N.: The original drawings are clearer than this (internet-)figure; for the original clarity, see the original work.)
Rothwell und Scheckler 1988, S. 86. (C. H. Beck, ed.: Origin and Evolution of Gymnosperms.)
Abb. 11
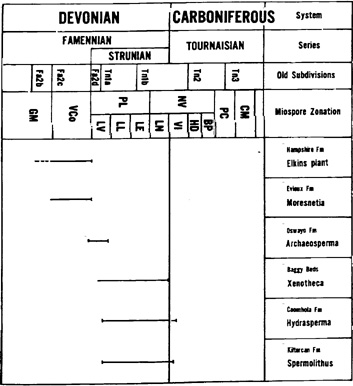
Figure 2.1. Stratigraphic chart showing the occurrence of Upper Devonian gymnosperms and their correlations with the European spore zonation. (Modified from Fairon-Demaret 1986).
(N.: The original drawing is much clearer than this (internet-)figure; for the original clarity, see the original work.)
Die hier sowie auf der folgenden Seite wiedergegebenen stratigraphischen Karten von oberdevonischen Gymnospermen verdeutlichen zum einen noch einmal die Tatsache, dass schon lange vor Genomosperma zahlreiche Samenanlagen von verschiedenen gymnospermen Gattungen bekannt sind und zum anderen, dass alle Gattungen 'sprunghaft' in Erscheinung treten und nicht durch viele nachgewiesene kleine Schritte voneinander oder von früheren Formen ableitbar sind. Gensel und Andrews bemerken 1984, S. 282 (Plant Life in the Devonian):
"Taking the definition of an ovule (= seed in this discussion) as 'an integumented megasporangium' the most primitive seed that we know about is Genomosperma kidstoni in which the megasporangium is only 'loosely' enclosed by sterile integumentary filaments. Most botanists seem to agree that this kind of structure was arrived at through a chain of development beginning with plants such as the homosporous rhyniophytes with a terminally borne single sporangium passing through stages in heterospory in which the number of megaspores was ultimately reduced to one. We have no positive information on such a chain of development as far as Genomosperma itself is concerned."
Für die 'Elkins'-Pflanzen fehlen die Übergangsformen genauso.
Die Samenanlagen dieser älteren Formen sind z. T. komplexer als die einiger Genomospermen. Wie oben zitiert, sind Rothwell und Scheckler aufgrund vergleichender Studien verschiedener ontogenetischer Stadien der Samenanlagen zu dem Schluss gekommen, dass die unterschiedlichen Grade der Verwachsung bei den 'Elkins'-Pflanzen eher ontogenetisch als phylogenetisch zu deuten sind. Könnte das nicht auch auf Genomosperma und Eurystoma zutreffen? Deutet man die Befunde jedoch genetisch und phylogenetisch im Sinne einer 'Höherentwicklung', so muss man annehmen, dass die Verwachsung von 'Hülltelomen' zum ersten Integument mehrmals
Abb. 12
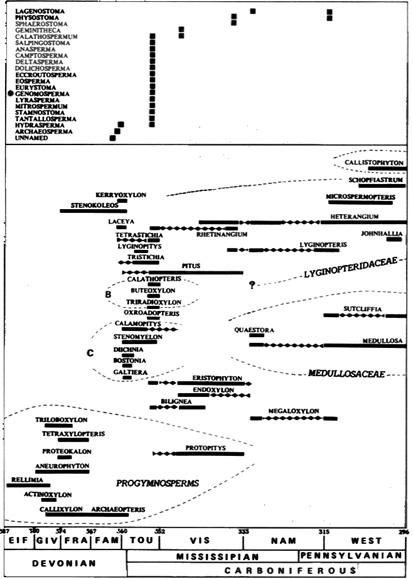
J. Galtier 1988, S. 138 (C. H. Beck, ed.):
Figure 3.1. Stratigraphical range of some early gymnosperm genera represented by permineralized sterns (below) and ovules (above), and comparison with a range of selected Devonian progymnosperms and Pennsylvanian pteridosperms of the families Lyginopteridaceae, Medullosaceae, and Callistophytaceae. B and C Genera attributed to the families Buteoxylonaceae and Calamopityaceae. The distribution of progymnosperms and of Pennsylvanian pteridosperms is based respectively on data from Beck (1976) and from Phillips (1981). Unpublished data included in this figure are kindly acknowledged-e.g., the extension of Lyginopteris down into the Upper Visean (U. Bertram, personal communication), the description of the new calamopityan genus Galtiera (Beck and Stein 1987), and the discovery of Lagenostoma in Upper Visean-Lower Namurian of Morocco (Galtier, Phillips and Chalot-Prat 1986).
(N.: The original drawing is much clearer than this (internet-)figure; for the details and original clarity, see the original work.)
unabhängig voneinander bei den verschiedensten Gattungen und z. T. in Abständen von Jahrmillionen erfolgt ist. Da die unterschiedlichen Grade der Verwachsung jedoch sowohl bei den ältesten bisher gefundenen Samenanlagen der 'Elkins'-Fossilien als auch bei den später auftretenden Genomospermen jeweils etwa gleich alt sind und auch nach einer etwa 10 Millionen Jahre währenden Selektion (vom Famennian bis ins Unterkarbon) noch existierten, ist eine weitere evolutionistische Deutung möglich: Die Verwachsung ist bei einigen Formen sekundär unterblieben. Zur Veranschaulichung eines solchen Degenerationsprozesses könnte man viele Mutanten aus der Gegenwart aufführen, die zugleich die Notwendigkeit einer adäquaten komplexen genetischen Grundlage für viele 'Verwachsungsprozesse' veranschaulichen würden. Jedenfalls stellt sich die Frage nach der genetischen Grundlage der Verwachsung von Hülltelomen zum ersten Integument (welches ich jedoch aufgrund einer mindestens 150 Millionen Jahre währenden Kluft zwischen den Angiospermen und dem hypothetischen Anschluss u. a. an die Lyginopteridopsida nicht ohne weiteres für das (erste) Vorläuferintegument der Blütenpflanzen halte).
Was die verschiedenen Deutungsmöglichkeiten der unterschiedlichen Verwachsungsgrade anlangt, so scheint mir die funktionell-ontogentische Interpretation von Rothwell und Scheckler bisher am besten abgesichert zu sein. Das würde jedoch schon eine 'komplexe Genetik' voraussetzen, wie das ja übrigens auch für weitere hochdifferenzierte anatomische Zusatzbildungen devonischer und karbonischer Samenanlagen (ich denke besonders an die Salpinx) zu fordern ist. Das im Laufe der Ontogenese für verschiedene Verwachsungsgrade codierende genetische System könnte dann auch mutieren und u. a. heterochronische shifts verursachen, so dass - neben sekundär offenen Samenanlagen - auch eine wiederholte frühe Verwachsung der 'Hülltelome' zu unterschiedlichen Zeitperioden denkbar wäre. Den Ursprung des variablen Systems hätten wir damit jedoch weder auf der genetischen noch auf der anatomischen Ebene erfasst.
Auch das fast gleichzeitige Auftreten vieler unterschiedlicher Formen ist mit der Idee der kontinuierlichen Entwicklung schwer in Einklang zu bringen (Abb. aus Strasburger 1991, S. 697). (Allerdings müssten inzwischen einige Linien leicht nach unten korrigiert werden, was jedoch nichts an der grundsätzlichen Situation ändert.)
Die Entfaltung der wichtigsten Pflanzengruppen während der Erdgeschichte. (Nach Mägdefrau.
(N.: The original drawing is much clearer than this (internet-)figure; for the original clarity, see the original work.)
Abb. 13
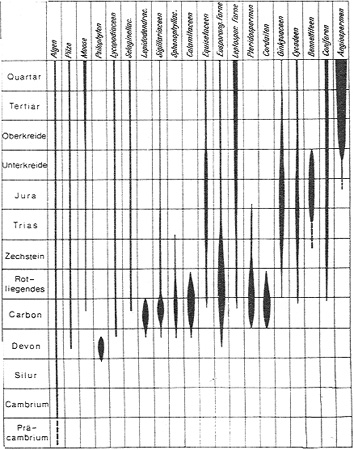
"By the late Devonian all extant major plant groups, except the Angiosperms, were recognisable" (Thomas and Spicer 1987, S. 73).
Mit ein paar Stammbäumen, die noch einmal den generellen Trend des diskontinuierlichen Auftretens neuer Formen in der Paläobotanik veranschaulichen, möchte ich die Ausführungen zur Frage nach der Entwicklung des Pflanzenreichs vorerst einmal abschließen ( obwohl mich eine systematische Behandlung der Thematik sehr "reizen" würde - aber dafür fehlt mir die Zeit).
Abb. 14
Stammbaum aus Strasburger 1991, S. 712.(
(N.: The original drawing is of a much better quality than this (internet-)figure; for the original clarity, see the original work.)
Hätten wir nur viele kleine Schritte, wie sie auch beim Vergleich verwandter Arten in Erscheinung treten, dann brauchten wir (zumindest bei den fossil gut überlieferten Formen) die stammesgeschichtlichen Zusammenhänge nicht zu vermuten, sondern könnten die Entwicklung im Detail aufzeigen. Das ist jedoch weder zwischen den Gruppen der hier aufgeführten Samenpflanzen, noch generell innerhalb der Gruppen der Fall. Da das auch auf die mit Millionen von Funden der am besten überlieferten Tiergruppen zutrifft (vgl. Zitat Kuhn oben), erhebt sich die Frage, ob man nicht einen ganz anderen Modus der Entstehung der Lebensformen als den der kleinen Schritte postulieren sollte - einen diskontinuierlichen Modus, der mit den paläobiologischen Befunden im Einklang ist.
Abb. 15
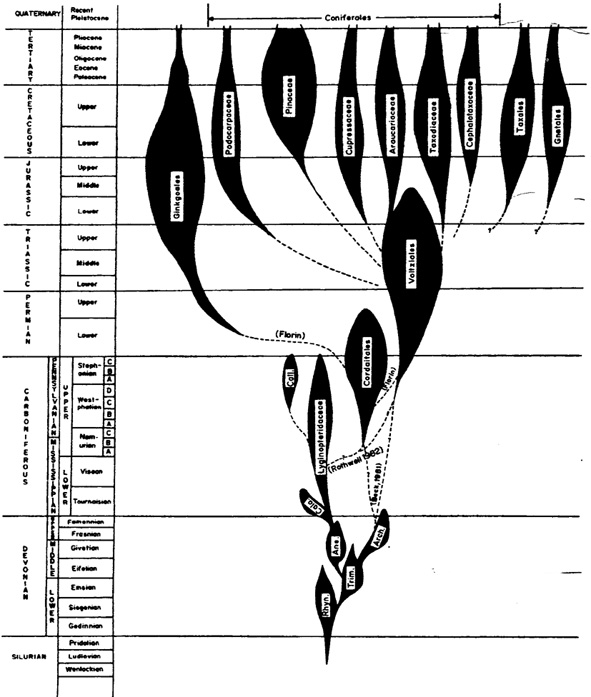
Stammbaum aus Stewart and Rothwell 1993, S. 420:
"Chart 29.1. Suggested origins and re1ationships of Ginkgoales, Gnetales, and major groups of coniferophytes, and their distribution in geologica1 time. Rhyn. = Rhyniopsida; Trim. = Trimerophytopsida: Ane. = Aneurophytales; Cala. = Calamopityaceae; Arch. = Archaeopteridales; Call. = Callistophytaceae
(N.: The original drawing is much clearer than this (internet-)figure; for the original clarity, see the original work.)
Bei den durchgezogenen Linien unterscheiden die Autoren nicht mehr zwischen ihren Vermutungen und den Realitäten. Interessanterweise leiten Stewart und Rothwell die Archaeopteridales nicht von den Aneurophytales, sondern beide unabhängig von den Trimerophytopsida ab, -womit die geniale Erfindung des bifazialen, sekundäres Xylem und sekundäres Phloem bildenden Cambiums zweimal unabhängig voneinander durch viele kleine Schritte (richtungslose 'Mikromutationen') erfolgt wäre. Die z. T. auch in der Abbildung wiedergegebenen widersprüchlichen Ableitungen verschiedener Gruppen zeigt wieder, dass das postulierte Kontinuum mit vielen kleinen Schritten fossil nicht belegt (nicht "zu sehen") ist, sondern zur Zeit nur "geglaubt" werden kann. Leitet beispielsweise C. die Gingopsida von den Progymnospermae ab, so favorisieren Stewart und Rothwell mit Florin die Ableitung der Gingkogewächse von den Cordaitales. Die Pinaceae werden von Stewart und Rothwell von den Voltziales abgeleitet, von C. von den 'Cordaitidae' usw.
Abb. 16
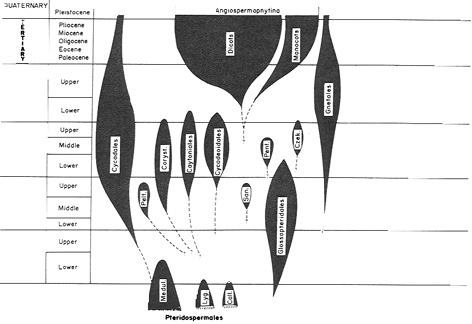
Stewart und Rothwell 1993, S. 444:
"Chart 30.1. Summary of suggested origins and relationships of angiosperms, cycadophytes, and glossopterids and their distribution in geological time. Medul. = Medullosaceae; Lyg. = Lyginopteridaceae; Call. = Callistophytaceae; Pen. = Pentoxylales; Czek. = Czekanowskiales: San. = Sanmiguelia; Coryst. = Corystospermaceae; Pelt = Peltaspermaceae."
(N.: The original drawing is much clearer than this (internet-)figure; for the original clarity, see the original work.)
Auch hier zeigen sich beim Vergleich mit Strasburger Stammbaum wieder zahlreiche Diskrepanzen, die allesamt letztlich wieder darauf beruhen, dass eine kontinuierliche Entwicklung in vielen kleinen Schritten nicht nachweisbar ist. Mit zahlreichen weiteren Stammbäumen anderer Forscher könnte man wohl ein ganzes Buch mit detaillierten Diskussionen zu widersprüchlichen Ableitungen füllen, die diese Aussage nur weiter bestätigen würden. Mir erscheint daher der oben zitierte Ausspruch Salthes nicht unberechtigt.
Abb. 17
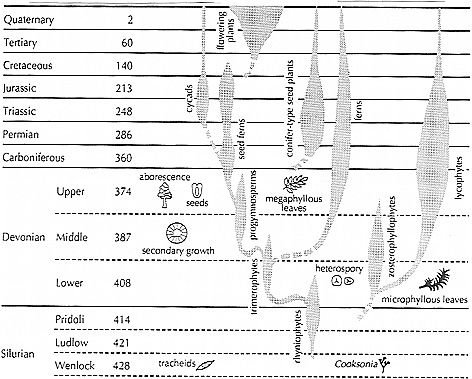
Stammbaum aus L. Graham 1993, S. 5 (Origin of Land Plants):
(N.: The original drawing is much clearer than this (internet-)figure; for the original clarity, see the original work.)
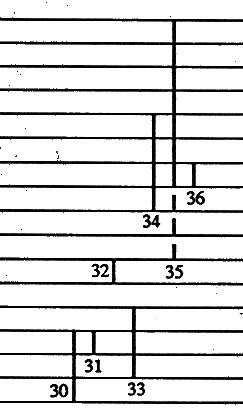
Die einzige mir bisher bekannte voll realistische Darstellung der fossilen Überlieferung auf neuerem Stand findet der Leser bei Benton (1993): THE FOSSIL RECORD (nur Wiedergabe der Progymnospermen: 30. Aneurophytaceae. 31. Protokalonaceae. 32. Protopityaceae. 33. Archaeopteridaceae. 34. Noeggerathiaceae. 35. Tingiostachyaceae. 36 Cecropsidaceae):
(N.: The original drawing is clearer than this (internet-)figure; for the original clarity, see the original work.)
Abb. 18