LAMPRECHTS KONZEPT DER INTRA- UND INTERSPEZIFISCHEN GENE
Sieht man sich Lamprechts Artdefinition näher an, so erhält man zunächst den Eindruck, dass es sich um eine Weiterführung des de Vriesschen Ansatzes handelt: Wenn Formen, welche bei reziproken Kreuzungen in allen Merkmalen den mendelschen Regeln folgen, als Varietäten derselben Art aufzufassen sind, dann lautet der Umkehrschluss, dass Formen, die nicht in allen Merkmalen diesen Regeln folgen, auch nicht zur selben Art gehören. Nun haben wir ja schon einige Anmerkungen zum undifferenzierten Umkehrschluss gemacht. Aber Lamprecht möchte sehr klar zwischen primären und sekundären Artbarrieren unterscheiden, wenn er schreibt (vgl. auch pp. 45/46 oben):
Handelt es sich nur um als Arten aufgefaßte Rassen, so bekundet sich dies in Kreuzungen zwischen solchen dadurch, daß die als artentrennend aufgefaßten Merkmale von der einen Art in die andere und vice versa überführt und in fertilen Nachkommen erhalten werden können. Ob die Kreuzung selbst normal fertil oder partiell steril ist, spielt hierbei keine Rolle, da eine teilweise Sterilität solchenfalls durch mehr oder weniger abweichende Chromosomenstruktur der Eltern bedingt sein kann Die scheinbar artentrennenden Merkrnale können hierbei auch kombiniert mit verschiedener Chromosomenstruktur in fertilen Individuen erhalten werden. Langjährige Studien haben dargetan, daß verschiedene Chromosomenstruktur allein, d. h. ohne verschiedenem Inhalt an artentrennenden Genen, keine unüberbrückbare Artbarriere bilden können, eine Auftassung, die auch GOLDSCHMIDT (1948) zur seinen gemacht hat
Lamprecht hat in jahrzehntelangen experimentalgenetischen Studien (vor allem an Pisum) im Detail gezeigt, 'wie man scheinbar artentrennende Merkmale...auch kombiniert mit verschiedener Chromosomenstruktur in fertilen Individuen erhalten' kann. Alle durch Chromosomenmutationen bedingte Sterilitätsgrade entfallen für den Artbegriff ebenso wie negative intraspezifische Geninteraktionen. Schon 1949 p. 22 schreibt er zum letzteren Punkt:
Es gibt auch binnen der Art genotypische Konstitutionen, die hohe Avitalität bedingen. Solchen werden dann natürlich ebenso und in noch höherem Grad interspezifisch anzutreffen sein. Aber die hierdurch verursachten Störungen...sind durch ungünstige Kombinationen der Allele von intraspezifischen Genen bedingt und haben mit der Barriere zwischen primären Arten selbst nichts zu tun.
Mit zahlreichen experimentalgenetischen Ergebnissen hat Lamprecht diese Schlussfolgerung untermauert (vergleiche die zusammenfassenden Arbeiten von 1966 und 1974).
Durch Genommutationen entstandene Arten werden als "sekundäre Arten" bezeichnet (z.B. 1944, 1966). Diese durch Genommutationen bedingte additive Artbildung erhöht zwar die Variationsbreite, sie gibt jedoch "keinen Aufschluß über die Evolution selbst, die zur Entstehung von "Arten primären Ursprungs", zu "primären Arten" führt" (1949, p. 3).
Nach Lamprecht haben die uns bekannten Gen-, Chromosomen- und Genommutationen (in welcher Kombination auch immer) mit dem Ursprung primärer Arten nichts zu tun. Abgesehen von unserer Diskussion dieser Fragen oben, stimmen ihm weitere Autoren in mehreren Punkten zu. So gibt beispielsweise Wagner (1983, p. 427) zu Grants Buch PLANT SPECIATION unter anderem zu bedenken:
There seems to be much more current interest in all phylogenetic characters and their significance than just in breeding systems and chromosome numbers. There is more and more emphasis on character divergence and shared divergent advancements as the essence of species diversification. True divergence in terms of many characters - geographical distribution, habitat, phenology, structure, physiology, chemistry - all participate in the process of speciation as well as our recognition and interpretation of taxa. In the absence of substancial divergence over the broad biological spectrum such phenomena as polyploidy, aneuploidy, sexuality, and crossibility may actually be trivial, mere "evolutionary noise", things that plants do that really don't amount to much in the big picture of speciation. Ordinary diploid, sexual, outbreeding populations shall inherit the earth; it is they that supply the fundamental divergences from the groundplans of their ancestors and that underlie plant speciation, not the polyploids, the apomicts, or the hybrids. The pillars of significant speciation are character polarities and character-state correlations as they bear upon the successful adaptations of divergent plant populations, grades and clades.
Da Wagner jedoch nicht die Möglichkeit einer Artabgrenzung durch Hybridisation von diploiden und sexuell sich fortpflanzenden Populationen ins Auge fasst (meiotisch-plasmatische Artbarriere), würde Lamprecht ihm bei der uneingeschränkten Vernachlässigung des Themas "breeding behavior" klar widersprechen.
Bleiben wir noch einen Augenblick beim Punkt "evolutionary noise": Stebbins diskutiert die Frage nach der Bedeutung der Polyploidie unter der Überschrift "Polyploidy and the Origin of Higher Categories" (1971, p. 199):
Cytologists have often debated the question: Does polyploidy produce only new variations on old themes, or can it be responsible for major evolutionary advances? The principal argument in favour of the latter point of view has been the existence of entire tribes, subfamilies or families (...Kapitelnachweis), which have basic numbers of obvious polyploid derivation. Evidence favouring the former point of view is that most of the more advanced families of flowering plants, such as Compositae and Gramineae, contain many genera having basic numbers which are not of polyploid origin. Where genera with high basic numbers exist, they are most easily explained as the polyploid derivatives of genera having low basic numbers which already possessed the morphological specializations of the family in question. More recently, the case against polyploidy as a progressive agent has been strengthened by the increasing number of chromosome counts in woody genera of various families which are so low that they cannot be of polyploid origin. There is now good reason to believe that when chromosome numbers of the majority of tropical woody genera become known, the phylogenetic sequences or families which include genera having original diploid basic numbers will be nearly as complete as those based upon the morphology of existing forms. The existence of such sequences would indicate that polyploidy has been important in the diversification of genera and species within families, but not in the origin of the families and orders themselves.
(Zu weiteren Überlegungen des Verfassers, vergleiche man das zitierte Kapitel.)
Stebbins schreibt in ähnlicher Weise 1980, p. 165:
Polyploidie ist ein sehr häufiger Weg in der Evolution der höheren Pflanzen; zwischen einem Viertel und einem Drittel aller Blütenpflanzen sind polyploid im Verhältnis zu ihren nächsten Verwandten. Bekannte Beispiele unter den Kulturpflanzen sind Weizen, Hafer, Kartoffel, Tabak, Baumwolle, Luzerne und die meisten Arten der Weidegräser. Bekannte polyploide Unkräuter und wilde Blumen sind Iris versicolor, Thalictrum ssp., einige Wildrosenarten, Stellaria media, Montia perfoliata, Achillea ssp. und verschiedene Arten von Veilchen und Astern. Darüber hinaus gibt es Hinweise, daß vor langer Zeit Gattungen und Gruppen von Gattungen, wie Apfel, Olive, Weide, Pappel und viele Farne durch Polyploidie entstanden sind.
Trotz allem hat Polyploidie wenig zum Fortschritt der Evolution beigetragen. In Gattungen, in denen Diploide neben Polyploiden vorkommen, werden alle Haupttrends der Evolution durch diploide Arten vertreten. Die Polyploiden dienen nur dazu, die Variationen gewisser besonderer adaptiver "Themen" zu vermehren. Der Grund hierfür liegt wahrscheinlich darin, daß die große Menge der Genverdopplungen die Wirkungen der neuen Mutationen und Genkombinationen in solchem Maße verdünnen, daß die Polyploiden große Schwierigkeiten haben, tatsächlich neue adaptive Genkomplexe zu entwickeln.
Beim Thema 'Genmutationen' stimmen den Lamprechtschen Ausführungen ebenfalls viele Autoren zu (z.B. Nagel 1980, pp. 170/171; Nes und Nes 1980, p. 194; Gitt (Hrsg.) 1982; Kahle 1984 u.v.a.).
Worin besteht nun nach Lanprecht die primäre Artbarriere? Im obigen Zitat (p. 210) klingt der Punkt schon an: "Mit Hinblick auf die unüberbrückbare Artbarriere kann kurz festgestellt werden, daß die für die Ausbildung von spezieseigenen Merkmalen des väterlichen Elters verantwortlichen Allele zusammen mit Homozygotie und Fertilität - im mütterlichen Plasma nicht reproduziert werden können (s.u.)." "...die intraspezifischen Gene sind verantwortlich für die genisch bedingte Variation der Merkmale innerhalb der Spezies. Die interspezifischen Gene sind verantwortlich für die Ausbildung von Merkmalen, die Spezies und höhere Kategorien trennen."
1966, p. 33 schreibt derselbe Autor:
Die unüberbrückbare Barriere zwischen naturbedingten Arten kommt nicht im Sterilitätsgrad des Bastards zum Ausdruck, sondern in der Nichtüberführbarkeit der arttrennenden Merkmale, die durch die Allele von interspezifischen Genen bedingt werden (s. L. 1948 b).
Lamprecht betont in seinen Ausführungen immer wieder die Korrelation zwischen Genom und Plasmon, ein Punkt, der von der Synthetischen Evolutionstheorie als unbedeutend für die Frage nach dem Ursprung der Arten zurückgewiesen wurde (vgl. z.B. Lüers, Sperling, Wolff 1974; Mayr 1967 und schon vorsichtiger 1982). In vielen Arbeiten wird die Frage nach der Bedeutung der plasmatischen Konstitution und die Korrelation mit dem Genom für den Artbegriff und den Ursprung der Arten nicht einmal gestellt (vgl. Erben 1975, Dobzhansky et al. 1977, Grant 1981, Willmann 1985 u.v.a.).
Nichtsdestoweniger erscheint die oben im Detail zitierte Spaltung 1 : 2 : 0 so einfach, klar und bedeutungsvoll, dass sich die Frage nach der Berechtigung dieses Ansatzes stellt.
Mehrere ältere Autoren haben sich mit dem [Genom-Plasmon-]Fragenkomplex beschäftigt (Correns 1901; Bateson 1909, 1924; Wilson 1925; Morgan 1927; Michaelis 1939, 1966 u.a.). Correns hat schon 1901 die Frage nach der Funktion des Plasmas gestellt. Beim Versuch für die 'Mischbarkeit der Erbanlagen auf der einen und die Stetigkeit in der Entwicklungsfolge auf der anderen Seite' eine Erklärung zu geben, schreibt er p. 87:
Bei dem Bestreben, den auf der Hand liegenden Widerspruch zu beseitigen, bin ich auf eine Vorstellung gekommen, mit der ich nicht hinter dem Berge halten will, obschon ich weiss, dass sie als arge Ketzerei aufgenommen werden wird. Ich möchte nämlich den Sitz der Anlagen, ohne feste Bindung, in den Kern, speciell die Chromosomen verlegen, und daneben noch ausserhalb des Kernes, im Protoplasma, einen Mechanismus annehmen, der für ihre Entfaltung sorgt. Die Anlagen können nun beliebig durcheinander gewürfelt werden, wie die bunten Steinchen in einem Kaleidoskop; sie entfalten sich an der richtigen Stelle.
Eine Consequenz dieser Ansicht ist, dass der Entfaltungsmechanismus beim Kinde im Wesentlichen der der Mutter sein wird, eine Annahme, vor der ich nicht zurückschrecke.
- Worauf unter anderem der Hinweis folgt, dass Driesch schon 1894 zu diesem Schluss kam.
Bateson haben wir oben (p. 209) schon zitiert. Er hat die Problemstellung präzis definiert. Wilson und Morgan haben aus entwicklungsphysiologischer Sicht die Bedeutung der cytoplasmatischen Konstitution des Eies für die Ontogenese hervorgehoben, und Michaelis ist dem Fragenkomplex in jahrzehntelangen experimentalgenetischen Studien auf den Grund gegangen
Der Anschluss an die älteren Autoren ist heute vor allem durch die moderne biochemisch-molekularbiologisch arbeitende Entwicklungsphysiologie gegeben. Im folgenden wollen wir eine Serie von Zitaten aufführen, in denen die Bedeutung des Plasmas hervorgehoben und begründet wird:
Michaelis 1966, p. 1:
Forty years ago, the question of the existence of cytoplasmic inheritance was debated. Therefore the first task was to find the irrefutable proof of the existence of cytoplasmic heredity and to refute all possible objections. Today the existence of cytoplasmic inheritance is generally acknowledged. There have been found many examples of cytoplasmic inheritance in various objects.
Ayala und Kiger 1980, p. 459:
Maternal-effect mutations demonstrate that development cannot proceed normally in the absence of a properly prepared egg cytoplasm, produced during oogenesis under control of the maternal genome. (Der hier angesprochene Kontrollmechanismus wird jedoch von vielen Entwicklungsphysiologen in dieser Ausschließlichkeit bestritten. Neuere Befunde wie Transkriptionsfaktoren sprechen hingegen z.T. wieder dafür.)
In seiner Arbeit 'Genetic and Developmental Studies of a New Grandchildless Mutant of Drosophila melanogaster' stellt Mariol 1981, p. 505 unter anderem fest:
...The death of embryos is maternally determined and hatching probability of an embryo does not depend on its own genotype.
(p. 510:) The identification of maternal determinants, and their role in development is, at the present time, one of the most exciting problems in biology.
Brinkley 1982, p. 1028 (Zusammenfassung aus: Cold Spring Harbor Symposia on Quantitative Biology, VOL XLVI: Organization of the Cytoplasm):
The organization of cytoplasm is far more complex than most investigators had imagined. Our view of the cytoplasm has progressed from that of a formless broth bathing the organelle to one of a dynamic interacting latticework of tubules, filaments and connections forming complex gel-sol transitions.
(p. 1029:) Indeed, as shown at this meeting, advances in technology and methodology have convincingly shown that the cytoplasm is highly organized at all levels. Cellular components as diverse as ions, amino acids, and enzymes are rarely if ever free in solution as one believed. They are instead a part of an ordered structural hierarchy. Even intracellular water (Clegg) appears to be largely bound to the cell matrix and therefore highly organized within the cytoplasm. Thus, the foremost principle of cytoplasmic organization appears to be that of order, spatial and functional order.
Hamburgh 1971, p. 138:
The evidence for feedback of cytoplasm on nuclei, so carefully put together by Briggs, King, Moore, Fischberg, Gurdon and Kroeger, is overwhelming and from it we have learned that nuclei behave and synthesize in accordance with the cytoplasm in which they find themselves.
(p. 115:) The nuclear transplantation experiments provided the first systematic evidence that the existence of a nuclear-cytoplasmic feedback is indeed a fact. Nuclear genes do not exist immutably and unchangeably behind the protective cover of the nuclear membrane, like the printer's plates behind glass, but there is a very lively interaction between nucleus and cytoplasmic environment leading to progressive change in the makeup of a cell. Moore has shown, as mentioned above, that blastula nuclei exposed to the cytoplasm of eggs of a different species are permanently changed, so that they became incapable of promoting normal development even after they have been returned to their own familiar cytoplasm. But the most dramatic evidence of the cytoplasmic environment on the nucleus came from studies by Gurdon and Brown. ...They could show that the nuclei derived from a population of highly differentiated intestinal cells that are actively engaged in ribosomal RNA synthesis, after transplantation into egg cells abruptly stop ribosomal RNA synthesis in the new cytoplasmic environment. Within 40 minutes after successful transplantation, nucleoli disappear from such transplant nuclei.
(p. 117:) Additional observations that can be interpreted in terms of direct intervention by cytoplasmic environment on gene activity during development were recently reported by Briggs and Justus in connection with the gene mutation o in the axolotl. ...this cytoplasmic factor is probably a gene product of the normal allele which might exert its corrective action through intervention on nuclear RNA synthesis.
(p. 125:) When salivary gland chromosomes were transplanted to preblastoderm and blastoderm cytoplasm new puffs appeared at chromomeric regions that had never exhibited swelling in the salivary cells before, while other puffs characteristic of salivary gland chromosomes rapidly disappeared.
(pp. 126/127:) ...a changing cytoplasm feeds back on the nucleus to trigger further differential gene activation.
Brothers 1981, p. 59:
The nature of the control of gene expression during development is one of the central problems in Developmental Biology. The egg, a giant specialized cell, apparantly contains morphogenetic components which are synthesized during oogenesis and stored in the oocyte. These components are distributed in a pattern which after fertilization acts to determine the developmental pattern for the main organ forming areas of the embryo (Morgan 1927). Normal development involves the interaction of these morphogenetic components and the embryo nuclei. This nucleo-cytoplasmatic interaction has long been thought to be at the basis of cell type determination.
De Robertis und Black 1981, p. 49:
When an egg develops all the cells inherit genetically identical nuclei, but nevertheless give rise to many distinct tissues. When analysing development, it is hard to avoid the thought that the initial differences between cells in an embryo must lie in the type of cytoplasm they inherit. (Vgl. auch Malacinski und Spieth 1979 p. 127.)
Cohen 1979, p. 11:
...to understand the first steps of morphogenesis we must understand not only the code but also the decoding machine.
Dohmen und Verdonk 1979, p. 127:
In most animal species the early stages of development are determined to a large extent by maternal factors that are synthesized during oogenesis. These factors are generally located in the cytoplasm or the cortex of the egg, and they are segregated into special regions of the egg by a process called 'ooplasmic segregation' (Costello, 1948) or 'cytoplasmic localisation' (Wilson, 1929). (Vgl. auch Smith und Williams 1979, p. 167 ff.)
Hadorn 1981, p. 22 (nach Aufführung mehrerer Beispiele von Bastardmerogonen):
Solche Erfahrungen zeigen nochmals, daß der Kern keineswegs die Entwicklung souverän beherrscht. Seine Gene sind vielmehr auf die Mitarbeit des Plasmas angewiesen, das eben auch über artspezifische Qualitäten verfügt. Und wenn ein Bastardmerogon - je nach Kombination früher oder später - zum Entwicklungsstillstand kommt, so beruht dies darauf, daß Kern und Plasma sich nicht mehr "verstehen" und so an der Lösung der geforderten Aufgaben scheitern müssen.
(Auf der nächsten Seite folgt jedoch die Einschränkung, dass die Merogonieversuche leider nicht erlauben, zwischen einer kernbedingten Prädetermination und einer autonomen plasmatischen Vererbung zu unterscheiden, da es durchaus möglich ist, dass das Plasma der Eizelle durch mütterliche Kerngene "prädeterminiert" wird.)
Davidson 1976, p. 317:
Whatever the nature of the morphogenetic determinants in egg cytoplasm, they must be synthesized during oogenesis. ...Evidence from many systems suggests that when finally localized these (morphogenetic determinants) are fixed in the cortex or subcortical regions of the egg. In some cases this may mean the egg membrane itself.
There is no convincing evidence as to the molecular mechanism by which the localized determinants of the egg function, only that they exist. In many cases they seem to be responsible for large programs of differentiation, such as the whole set of functions required for the differentiation of primordial germ cells or coelomic mesoderm, or cephalic plate cells.
(pp. 44/45/47:) ...certain complex early processes do continue to be carried out in the total absence of nuclei. ...The enucleation experiments...have demonstrated that the division of the egg mass into cells separated by membranes, the most obvious function of the cleavage process, is dependent on maternal cytoplasmic factors rather than on the embryo genome. ...From these experiments the important conclusion was drawn that protein synthesis in early embryos occurs on preformed templates, i.e. maternal messenger RNA. (Details dazu u.a. bei Thomas et al. 1981 in ihrer Arbeit "Molecular Structure of Maternal RNA'.)
Gurdon 1974, p. 111 (aus der Zusammenfassung des Kapitels 'Gene transcription and the initiation of cell differentiation'):
Egg cytoplasm contains a range of components which are of major importance in development because they regulate independently the activities of nuclei and genes, and because they appear to be responsible for promoting different directions of specialization in early development.
Das Kapitel 4 'The cytoplasm in differentiation' von Macleans Buch THE DIFFERENTIATION OF CELLS (1977) enthält folgende Untertitel:
(1) Modification of nuclear gene activity by cytoplasmic molecules
(2) Pre-existing cytoplasmic structures which are necessary for the synthesis of new structures
(3) Heterocaryons and nuclear transplants
(a) The cytoplasm affects nuclear size, in terms of both volume and mass
(b) The cytoplasms affects the timing of DNA synthesis
(c) The cytoplasm affects chromosome condensation and mitosis
(d) The cytoplasm affects nuclear RNA synthesis
(e) The cytoplasm, in some cases, diverts the direction of differentiation
(4) Cytoplasmic DNA
(a) Mitochondria
(b) Chloroplasts
Jeder Punkt wird detailliert anhand experimenteller Daten begründet (vgl. pp. 103 - 133 der Arbeit).
Raff und Kaufman 1983, pp. 110/111:
If differential gene action is to be elicted by agents localized in the cytoplasm, there should be evidence that cytoplasmic components can indeed direct the course of nuclear function. This evidence is available from experiments in which nuclei from one type of cell are transplanted into a host cell of a different type. A striking example of this experimental approach, as conducted in J. B. Gurdon's laboratory, was the injection of adult frog brain nuclei into three types of recipient frog cells by Graham et al. and by Gurdon. Adult frog brain nuclei do not normally synthesize DNA or undergo mitosis. Nuclei from these cells were injected into (1) oocytes, which synthesize RNA but not DNA; (2) ovulated oocytes undergoing the completion of meiosis and possessing condensed chromosomes on meiotic spindles; and (3) eggs immediately after activation, which are synthesizing DNA but not RNA. In all cases the injected nuclei changed their activities to conform to those characteristic of nuclei of the recipient cell types. This included, for instance, the condensation of chromosomes and their association with spindles in brain nuclei injected into maturing oocytes, or the synthesis of DNA by brain nuclei injected into activated eggs. Because neither of these activities is characteristic of normal brain nuclei, the new activities of the injected nuclei were determined by the host cytoplasmas. Similar nuclear transplant experiments have also shown that the transcription of specific genes in the injected nuclei (those for ribosomal RNA) is subject to regulation by host cytoplasm.
The influence of the cytoplasm on the nucleus extends to the eliciting of specific patterns of mRNA synthesis. DeRobertis and Gurdon injected the nuclei of cells of the frog Xenopus, grown in tissue culture, into oocytes of the salamander Pleurodeles. By high-resolution, two-dimensional gel electrophoresis they were able to distinguish between the protein synthetic pattern characteristic of Xenopus from that of Pleurodeles, and that of Xenopus cultured cells from that of Xenopus oocytes. When nuclei from cultured Xenopus cells were injected into Pleurodeles oocytes the Xenopus oocyte pattern of proteins was synthesized, not the cultured cell pattern. The shift was prevented by use of the RNA synthesis inhibitor, alpha-amanitin. Thus, the effect of the oocyte cytoplasmic environment on the Xenopus nuclei was to inactivate the expression of one set of genes and to activate the expression of another set characteristic of oocytes.
(p. 112): The existence of transcriptional as well as posttranscriptional mechanisms for differential gene expression by the nucleus will make the task of understanding the controls of gene action ultimately more difficult, but the existence of these mechanisms does not change the basic embryological issue. The hypothesis, diagrammed in Figure 42, is that certain macromolecules localized in the cytoplasm and partitioned into some of the embryonic blastomeres come to elicit specific patterns of gene expression by the nuclei of these blastomeres. (Kursivschrift immer von den Verfassern.)
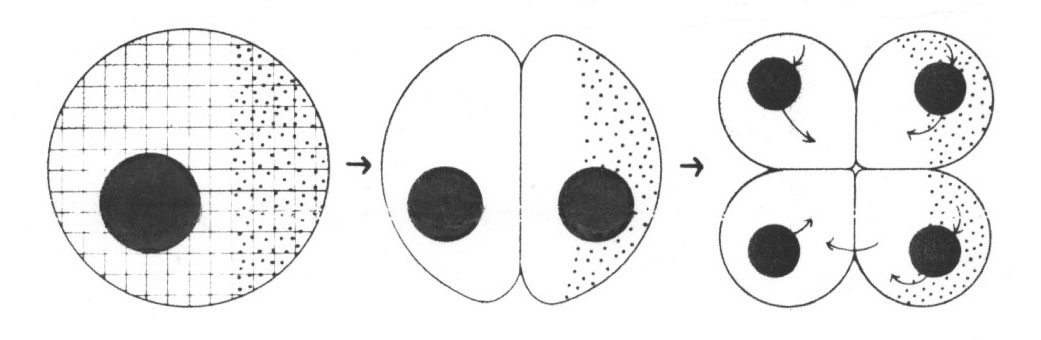
Abb. 42: Regionalized informational systems of eggs and embryos. The cytoskeleton is represented by a grid, the nuclei are back, and regionalized morphogenetic determinants are represented by stippling. Arrows represent information flow. The model is overly static, because in most cases morphogenetic determinants are nor prelocalized, but move to their final positions during the first few cleavages. (Aus Raff und Kaufmann 1983)
(p. 116): Perhaps because current fashion in molecular biology is so strongly intent on nucleic acids there has been less experimental emphasis on proteins as localized determinants of nuclear activity. Yet there is little question that such protein molecules exist.
Dillon postuliert aufgrund zahlreicher Befunde einen im Plasma gelegenen "supramolekularen genetischen Mechanismus", und er begründet seine Schlussfolgerung u.a. wie folgt (1983, pp. 434/435):
In seeking a mechanism that can induce, control, or carry out...changing gene activities, the most obvious one, the nature of the DNA, deserves the earliest attention. The single type of change that this gigantic macromolecule can effect is in its configuration, for as discussed in a previous section in all other processes it is the most inert of all cellular substances (Dillon, 1978a, p. 121). Thus, it is conceivable that the DNA molecule might be in a more tightly coiled configuration in transcriptionally inactive sectors that would unravel slightly when the developmental state was attained in which the particular sector was then needed. Since changes in pH or ionic content can modify the configuration of DNA, the concept appears to be sound - unless one attempts to determine how the requisite localized ionic or pH modulations are themselves induced. Then it becomes evident that an additional mechanism is required to govern the variations in the chemical properties from one section of the genome to another. Hence, the loosening or tightening of the molecular structure would itself not be a control mechanism but merely an effect of another less obvious one. Additionally this view is invalidated by those electron micrographs...cited and by numerous others of a similar nature. For it is readily apparent, as just pointed out, that the distance between nucleosomes is comparable in both the transcriptionally active and inactive regions, as would not be the case if unfolding the DNA backbone were involved. These same observations apply in like fashion to other methods of gene control that have been advocated, ...as changes in degree of methylation in active genes (for example, Naveh-Many and Cedar, 1981). Any such alteration does not represent the definitive mechanism, only an intermediate stage.
Consequently, the sole ultimate source of control must lie in the soma of the cell, that is, in the supramolecular genetic apparatus, as no other possible mechanism is in evidence. Phrased in different terms, the soma contains an organized system of proteinaceous, and perhaps other, substances that is somehow programmed to carry out selective reading of particular portions of the genome. The programming must be viewed not as a static arrangement but as sensitive to changes in the immediate environment of the cell and continually subject to modification by some of the products of the genes that are read. In brief, a feedback mechanism exists. At the earliest stages of embryogeny in which transcription occurs, the program is set to read a certain number of genes, especially the more ancestral types [nicht notwendigerweise "ancestral", Anm. von W.-E.L.], including one group that can be called protein set Q. The components of this set are not to be thought to be concentrated into one region of a certain chromosome but as a diversity of types from families of genes scattered throughout the genome. When the transcripts for this set have been translated, along with the others characteristic of this stage, the products effect a slight change in certain parts of the program, inducing the supramolecular genetic mechanism to read some new genes. The latter would not only encode more advanced replacements for previously synthesized substances, including protein set R, but would also code for additional products. The members of set R whose gene likewise would be scattered about the genome, would lead to another set of program changes, inducing the synthesis of protein set S, and so on throughout development.
Seine umfangreichen Untersuchungen zur Existenz und zu den Aktivitäten des "supramolekularen genetischen Mechanisnus" fasst Dillon auf den Seiten 444 - 446 wiederholt zusammen:
In summary, those cellular processes that seem to require the presence of an organized and programmed genetical mechanism outside the genome proper include the following that have been noted in the second and third volumes of this trilogy:
That these functions are conducted by an organized body within the soma of the cell, not merely by isolated independent enzyme systems, is evidenced particularly lucidly by the experiments on centrifuged fertilized ova reported in Chapter 3, Section 3.4.2. In spite of the strong centrifugation that displaced all internal organelles out of the typical animal-vegetal polar alignment, the treated cells regained the normal ovostratification as the embryo developed, oriented in normal relations to the typical axis.
Dillon 1987, p. 787 (nach Aufführung verschiedener Kontrollfunktionen; - siehe auch Fußnote unserer Arbeit p. 232):
Such a control probably involves a system of intricately coordinated proteins, possibly together with certain species of RNAs, that has been named the supramolecular, or second, genetic mechanism. But whatever name it finally receives, its existence is made undeniable by its numerous points of manifestation in gene regulation, in macromolecular behavior, somatic mutation, and throughout the cell cycle in organellar growth and function. (The Gene. New York.)
Gutmann und Bonik 1981, p.123:
Die Unterstellung, im Erbgut liege das Programm für den Organismus vor, beruht auf einem logischen Fehler. Wenn wir sicher nachweisen können, daß ohne Erbgut kein Lebewesen denkbar und ohne Erbgutveränderung keine Evolution möglich ist, so können wir den Schluß nicht umdrehen, aus Erbgut und Erbgutveränderungen alleine lasse sich die Erklärung für den Aufbau des Organismus und seine Leistungen gewinnen. Notwendige Bedingungen sind nur unter Verletzung der Logik in zulängliche umzudeuten. ...Wenn es also Vererbung gibt, dann kann sie nicht auf die Gene und Chromosomen als Informationsträger beschränkt sein. Es sind zur Verwirklichung eines Organismus komplexe Gefüge nötig, die selbst wieder Informationscharakter haben (plasmatische Vererbung).
Duspiva (1980, p. 11) (nach detaillierter Aufführung der Kerntransplantations-Untersuchungen von Gurdon - vgl. Zitat Raff und Kauffmann):
Aus diesen und vielen ähnlichen Versuchen kann der Schluß gezogen werden, daß es das Cytoplasma ist, welches die Kernaktivität steuert. Das Cytoplasma enthält in den betrachteten Fällen Induktoren für DNA- bzw. RNA-Synthese, die unabhängig voneinander funktionieren. Das gleiche gilt für die Repression dieser Funktionen.
...(pp. 11/12): Die Etablierung von Determinanten und ihre Auswirkung auf Entwicklungsprozesse können zeitlich weit auseinander liegen. Einen gewissen Einblick in die Wirkungsweise cytoplasmatischer Faktoren gibt folgendes Beispiel: Das Axolotl (Ambystoma mexicanum), ein neotänischer Molch, besitzt das Gen (0). Weibchen, die für das mutierte Gen (0-), ova deficient, homozygot sind, legen Eier, die sich nicht weiter als bis zum späten Blastulastadium entwickeln können, und zwar auch dann nicht, wenn sie mit Spermien befruchtet werden, welche das normale Gen (0+) mitbringen. Der Effekt von (0-) ist daher rein mütterlich. Die genetische Störung findet bereits im Ovar statt, wo in den Oocyten ein cytoplasmatischer Mangel entsteht, der sich erst viel später in einer Störung der Morphogenese auswirkt, wenn der Embryo in die Phase der Gastrulation eintritt. Diesen Entwicklungs-Stop kann man verhindern, wenn man in das mutante (0-)-Ei den Kemsaft oder einfach Cytoplasma einer reifen normalen (0+)-Oocyte injiziert (BRIGGS und CASSENS, 1966). Normale (0+)-Oozyten bilden während ihres Wachstumsstadiums, das durch "Lampenbürsten-Chromosomen" und hohe synthetische Leistung ausgezeichnet ist, eine 0+-Substanz. Wenn das stark vergrößerte Keimbläschen am Ende der Oogenese platzt, so tritt der Kernsaft aus. Mit ihm gelangt auch die 0+-Substanz in das Ooplasma. Dieser Faktor, vermutlich ein hochmolekulares saures Protein, verbleibt von diesem Moment ab im Cytoplasma der Eier und ist bis zum späten Blastulastadium nachweisbar. Dann verschwindet der Faktor; er wird entweder abgebaut oder an eine Struktur gebunden. Man weiß, daß sich der Amphibienkeim bis zum späten Blastulastadium ohne eine funktionelle Beteiligung des Genoms entwickelt. Von da ab beginnt das Erbgut des Keimes zu wirken. Um nun die Gastrulation in Gang zu bringen, ist die Transkription gewisser Abschnitte der DNA notwendig. Dieser Prozeß hängt aber von der Anwesenheit der 0+-Substanz ab (BRIGGS und JUSTUS, 1968; BRIGGS 1972). Mutante Blastulae bilden wenig oder gar keine RNA.
Nehmen wir in der 0+-Substanz ein während der Oogenese gebildetes Protein an (es könnte sich auch um eine stabile m-RNA handeln), so erhebt sich die Frage nach der Regulation des 0+-Gens: Welche Prozesse beschränken die Genaktivität auf die Oogenese? Erwarten wir mit den heutigen Genregulationsmodellen für Eukaryonten (vgl. z.B. Alberts et al. 1983) die Aktivierung durch ein Genregulations-Protein, dann stoßen wir auf das Problem der Regulation des Regulators. Neuere Befunde weisen wieder in Richtung Cytoplasma (Mitrofanow und Sidorova 1981, Graham und Wareing 1984, Jeffery 1985, Ingham 1985, Ghysen et al. 1985), welches zusammen mit den Kernstrukturen 'das vollendetste kybernetische System auf unserer Erde' bildet (so Struggers Kommentar zum Thema Zelle).
Die meisten Molekularbiologen neigen heute dazu, die Genregulation allein mit dem fast ausschließlich DNA-orientierten Konzept der differentiellen Genexpression zu erklären. Smolik-Utlaut gibt jedoch (1986, pp. 519-520) bei der Besprechung des Buches von De Pomerai (FROM GENE TO ANIMAL: AN INTRODUCTION TO THE MOLECULAR BIOLOGY OF ANIMAL DEVELOPMENT) unter dem Titel "A molecular biologist's view of development" folgendes zu bedenken:
Although differential gene expression is an important driving force in development, there are other, equally compelling forces at work. Morphogenetic gradients are a recurrent theme in developmental biology and could be discussed in more depth, perhaps using hydra as an example. Structural constraints are also critical to proper development. With this in mind, the work of J.G. Gerhart could be presented, his classic molecular experiments have demonstrated the importance of cytoplasmic rearrangements in establishing the dorsal-ventral axis of the Xenopus embryo.
Eine gute Zusammenfassung der Hauptpunkte findet sich auch bei Barry 1986.
Um die Problematik mit einem drastischen Beispiel zu veranschaulichen: Selbst wenn ein Planet von der Größe unserer Erde mit einer tausend Meter dicken, ausschließlich aus menschlichen Genomen bestehenden DNA-Schicht bedeckt wäre, so würde doch allein aus der DNA niemals ein Mensch entstehen.
Es bedarf der Integration der DNA-Information in ein (art-) spezifisches, lebendes System, um einen lebenden Organismus zu erzeugen. Reine DNA ist "tote Materie" (die zwar ungeheuer informationsreich sein kann, aber allein nicht lebendiger ist als etwa eine Enzyklopädie).
Blechschmidt beschreibt seine Auffassung zum Thema DNA und Differenzierung 1982, pp. 42 - 44, wie folgt (ich zitiere diese anregenden Gedanken, ohne in allen Punkten zu folgen):
Im Rahmen dieses Konzeptes ist die Frage, wie die materielle Beschaffenheit, die Chemie der Gene, mit der körperlichen Gestaltung in Beziehung steht, nach wie vor ungelöst. Es ist zwar sicher, daß die genetische Substanz beteiligt ist an der Eiweißbildung, es ist aber nichts darüber bekannt, wie die Eiweißbildung ihrerseits zur dreidimensionalen Gestaltung des Körpers beiträgt. Der Weg zur Verwirklichung des sogenannten Genotyps in den Phänotyp ist völlig ungeklärt. Das hängt u.a damit zusammen, daß es keine Gestaltungsstoffe, sondern nur Gestaltungskräfte gibt. Diese Gestaltungskräfte wiederum sind jedoch keine physikalischen Kräfte an sich. Den Begriff der Gestaltungskraft gibt es in der Physik nicht, worauf besonders Heitler hinweist. Gestaltungskräfte sind Ausdruck eines Gestaltungsprinzips, das auf der Grundlage des Lebendigen mehr ist als physikalisch-chemisch verrechenbar. Die chemischen Prozesse und die räumliche Gestaltung des Organischen sind zwei verschiedene Ordnungen, die unmittelbar nicht aufeinander rückführbar sind. Die DNS kann niemals eine Zelle machen; eine Zelle ist vielmehr schon zu Beginn der Differenzierung gegeben und hat bereits eine Gestalt, bvor sie zu wachsen beginnt und dabei Änderungen ihrer Gestalt hervorbringt. Zellkern und Zelleib sind nicht aufeinander zurückführbar. Dabei muß ein Stoffwechselkreislauf in der Zelle vorausgesetzt werden. Stoffaufnahme durch die Zellmembran, also von außen, führt über das Zellplasma zu Reaktionen im Kern und von dort rückwirkend zu Stoffwechselprozessen im Zytoplasma bis zur Zellgrenzmembran. In diesem Kreisprozeß spielt die genetische Substanz eine wichtige, aber für die Differenzierung nicht zureichende Rolle. Differenzierung verlangt danach mehr als genetische Information. Sie verlangt auch Gestaltungsarbeit eines zugehörigen extragenetischen Substrats. Hier sind viele komplexe Faktoren wie Wachstumsverschiedenheiten, Oberflächenspannungen etc. beteiligt. Darauf weisen besonders die Botaniker hin, die es mit vergleichsweise einfachen Objekten zu tun haben.
Die DNS könnte als ein Kochbuch aufgefaßt werden, das der Körper für seine Differenzierung braucht. Unklar ist aber, wer der Koch in der Küche ist. Unklar ist besonders, wie der Körper aus der Menge der bereitgestellten Rezepte auswählen kann, um die richtigen Gene zu benutzen. Denn die Existenz der Gene allein bedeutet noch nicht, daß sich aus ihnen der Verlauf der Differenzierung ergibt. Mit der Kenntnis von Genmolekülen ist die Frage, wie oder wann es geschieht, daß sich die Zellen differenzieren, noch nicht gelöst. Denn im Organismus sind alle Zellen mit den gleichen Genen ausgestattet.
...Gene sind eine unabdingbare, notwendige Voraussetzung, aber keine zureichende Bedingung für die Entwicklung und Differenzierung. Dies zu bemerken scheint wichtig, um nicht einer materialistischen Simplifizierung der Lebensvorgänge Vorschub zu leisten.
Es besteht kein Zweifel, dass die Gene eine große Bedeutung für die Vererbung und damit indirekt auch für die Differenzierung haben. Aber schon in dem jungen Keim sind weder die Chromosomen noch die Gene kinetisch aktiv. Sie haben keinen "Treibstoff" zu aktiver Tätigkeit. Sie sind nicht die Motoren der Entwicklung und bringen daher nicht selbst die Merkmale des differenzierten Organismus hervor. Auch nicht indirekt auf dem Weg über die von ihnen gebildeten Enzyme. Diese Annahme würde das Problem nur von der Chemie der Gene auf die Chemie der Enzyme verlagern. Selbst die wichtigsten Enzyme sind nur Beiträge, wenn auch unabdingbare Beiträge zu dem Prozeß der ganzen Differenzierung, der außer der genetischen "Information" immer noch andere Voraussetzungen hat. Die Zelle ist viel mehr als die Gene. Sie ist ein Ganzes (Zellkern, Zellplasma mit Organellen und Zellmembran). Und nur im Rahmen dieses Ganzen und als Teil dieses Ganzen haben die Gene Bedeutung.
Worin besteht nun aber der Beitrag der Gene? Die Gene sind chemische Konstanten des schon mit der Eizelle gegebenen individualspezifischen Stoffwechsels und als solche besonders stabile Bestandteile der Zelle. Sie legen wichtige materielle Voraussetzungen für den Stoffwechsel fest und informieren dadurch die wachsende Zelle gleichsam über ihre Möglichkeiten der Differenzierung. Und darin liegt ihre Bedeutung. Sicher ist, daß die genetische Substanz der Erhaltung der Individualität dient, während umgekehrt die extragenetische Substanz, vor allem das Zytoplasma, den Wechsel des Erscheinungsbildes während der Entwicklung gewährleistet. Weil Gene so besonders stabil sind, sind sie viel zu träge, um von sich aus zu agieren. Vielmehr antworten sie auf Reize vom Zytoplasma (von außen). Gene agieren nicht, sondern reagieren.
Brenner 1981, p. 4:
We may imagine someone standing up at some future conference to declare "I will now tell you everything about Drosophila' and then proceeding to intone 'AGGCTTA' and so on for 108 or so bases. For small viruses or for mitochondria, this would make sense, but for organisms of the complexity of Drosophila the sequence would be largely meaningless. The syntax would be there but the semantics would be absent.
Graham und Wareing 1984, pp. 5/6:
...the time of gene expression may be controlled at the translational level.
...the cytoplasm...plays a primary role in determining the time and place of expression of specific genes, i.e. it controls the selective gene expression underlying differentiation.
Zahlreiche weitere Autoren können zitiert werden (Jinks 1967, Markert und Ursprung 1971, Markert 1982, Sager 1972, Schwanitz 1974, Lovtrup 1974, 1977, P. von Sengbusch 1979, Korochkin 1980, Tsunewaki (Ed.) 1980, Luria, Gould und Singer 1981, Thomas et al. 1981, Scherrer 1981, Steward and Hunt 1982, Bonner (Hrsg.) 1982, Lima de Faria 1983, Kratzer 1983, Alberts et al. 1983, Lüning 1984, Nieuwkoop, Johnen und Albers 1985, Metz und Monroy 1985 und viele andere, die die Bedeutung der (Ei-)Plasmakonstitution für die Genregulation und Ontogenese hervorheben (einige Autoren werden wir später noch zitieren). Wie die Zitate zeigen, gibt es derzeit unter informierten Biologen nur noch Meinungsunterschiede zur Frage, ob von der im Kern gespeicherten Information unabhängig ein zweites, gleichwertiges (oder sogar übergeordnetes) Informations- und Regulationssystem im Plasma existiert oder ob letztlich alles oder zumindest der größte Teil aller wichtigen Informations- und Steuerungsprozesse vom Zellkern aus beherrscht und dirigiert werden. [Kaskaden von Regulatorgenfunktionen könnten hierfür noch zitiert werden.] Die zelluläre Struktur selbst bringt jedenfalls die Eizelle mit, wozu u.a. die (meisten) Mitochondrien und bei den Pflanzen die Plastiden gehören (eine recht umfangreiche Liste der 'maternal effects on development' hat Cohen 1979, pp. 26 - 28 vorgelegt). Wahrscheinlich ist der ganze Streit über diese Frage insofern müßig als die Zelle mit Kern, Plasma und Membranen ein ungeheuer komplexes kybernetisches System bildet, in welchem die integrierten Regelkreise eben alle drei Bereiche unabdinglich umfassen (vgl. Scherrer 1981, Steward und Hunt 1982, p. 51; im Prinzip schon Wilson 1925).
Für unsere Fragen nach dem Artbegriff und dem Ursprung der Arten entscheidend ist die Tatsache, dass nach allen bisherigen Ergebnissen und Erfahrungen feststeht, dass das Genom (DNA-Information) des Vaters mit dem plasmatischen Genregulations- und Expressionssystem der Mutter art-spezifisch übereinstimmen muss, um exprimiert werden zu können. Was heißt nun dieses "artspezifisch" molekulargenetisch und an welchem Punkt werden diese artspezifischen Grenzen erreicht oder überschritten und welche physiologischen und morphologen Konsequenzen sind bei primären Arten damit verbunden? Hier befindet sich das Problem der Artgrenzen und des Ursprungs neuer Arten.
Ohno hat sich sich dem Problem 1969 auf molekulargenetischer Ebene auseinandergesetzt und kommt zu ähnlichen Ergebnissen wie Lamprecht, wenn er p. 137 seine Ergebnisse wie folgt zusammenfasst:
We have studied embryonic development of rather extreme hybrids such as Rainbow trout female x Brown trout male, Japanese quail female x Button quail male, using the gene loci for various enzymes as markers. The results showed that in development of rather extreme hybrids, paternally derived alleles of many gene loci are ignored by the gene activation mechanism. As a result, these embryos go through the maternally hemizygous stage before and during organogenesis. Lethality of extreme hybrids early in development can now be explained on the basis that functionally they remain equivalents of the gynogenic haploid embryo. Two assumptions which are not mutually exclusive can explain preferential activation of maternally derived alleles in rather extreme hybrids; (1) the information which activates a group of structural loci very early in development is transcribed not by the nuclei of the zygote itself, but the diploid nucleus of the mother during oogenesis, and this information is stored in the egg cytoplasm before ovulation, (2) so far as the regulatory gene loci in the zygotic nucleus which control the activation of structural genes later in development are concerned, as a rule, only the maternally derived alleles function while corresponding alleles contained in the paternally derived genome (haploid set) remain dormant.
In either case, the gene activation mechanism which directs embryonic development is exclusively maternal. Thus, in the case of rather extreme interspecific hybrids, the gene activation mechanism of an embryo should encounter difficulty in recognizing paternal alleles of structural gene loci, as they are derived from an alien species.
(Details vgl. die Originalarbeit des Verfassers). Daraus folgt dann natürlich auch, dass Mutationen in Richtung 'artfremder Allele' Grenzen gesetzt sind und dass bei Überschreitung dieser Grenzen, solche Allele nicht mehr (oder nur noch ungenügend) exprimiert werden konnen - dazu vergleiche man die großen Kapitel von Lamprecht über seine Exmutanten von Pisum (1966, 1974).