WEITERE HINWEISE AUF EIN PLASMATISCHES REGULATIONSSYSTEM
Heitler diskutiert (1970, pp. 50/51) die Frage nach dem möglichen Informationsgehalt des Zellkerns im Vergleich zu der in hochkomplexen Lebensformen wie dem Menschen realisierten Information und schreibt:
Ob alle morphologischen Eigenschaften eines Lebewesens - die ja auch alle vererbt werden - sich in der Kombinatorik der Nukleotide widerspiegeln, ist sehr zweifelhaft. Der Kausalzusammenhang ist jedenfalls unklar. Die Zahl der Informationseinzelheiten, die durch die möglichen Anordnungen der Nukleotide in der DNS gespeichert werden kann, ist zwar unvorstellbar groß; trotzdem muß man zweifeln, ob diese Zahl ausreicht um alle morphologischen Einzelheiten des Körperbaus eines Organismus wiederzugeben.
Eine einfache Rechnung möge das illustrieren. In der DNS einer menschlichen Zelle sind - sagen wir - 5 Milliarden Nukleotide enthalten. Nehmen wir an, die 4 Sorten seien zu gleichen Teilen vertreten. Die Anzahl der Anordnungsmöglichkeiten in sämtlichen Chromosomen ist eine Zahl mit 3 Milliarden Nullen. Das menschliche Gehirn enthält 10 Milliarden Zellen, Neuronen genannt, die in der kompliziertesten Weise dureh Nervenstränge verbunden sind. Von jeder dieser Zelle gehen rund 100 Nervenstringe aus. Nehmen wir der Einfachheit halber an, es seien je zwei Neuronen paarweise verbunden. Die Zahl der Verbindungsmöglichkeiten allein ist schon eine Zahl mit 10 Milliarden Nullen. Und hierbei ist die Kompliziertheit des Gehirns stark untertrieben, und von anderen Körperteilen war gar keine Rede. Es ist also unmöglich, daß jede morphologische und anatomische Einzelheit in der DNS ausgedrückt ist. Eine kleine Änderung in der DNS muß schon größere anatomische oder morphologische Änderungen zur Folge haben.
Der morphologische Bau eines höheren Tiers ist im einzelnen derartig kompliziert, daß man kaum annehmen kann, daß die gesamte Information darüber in einigen, wenn auch noch so komplizierten Molekülen enthalten sein kann. Manche Biologen hahen auch die Ansicht vertreten, daß die vererbten morphologischen Eigenschaften ihren Sitz im Zellplasma hahen.
Eigen kommt (1983, p. 76) zum gleichen Schluss beim Versuch, die Vollmertschen Wahrscheinlichkeitsrechnungen zum Ursprung der Lebensformen zu widerlegen:
Es ist gar nicht möglich, daß der komplette Organisationsplan der Zellen im einzelnen in den Genen codiert ist; denn die Zahl der Zellen eines Säugetiers übersteigt um ein Vielfaches die Zahl seiner Gene, ja sogar die Zahl seiner Genbausteine. Das menschliche Gehirn zum Beispiel umfaßt allein etwa 10 Milliarden Nervenzellen, von denen jede wiederum durch etwa tausend bis zehntausend Kontakte (Synapsen) mit anderen Nervenzellen verschaltet ist. Diese Verschaltung ist organisiert, und die Existenz einer solchen Organisation ist die Grundlage der Funktion des Gehirns. Das menschliche Genom besitzt jedoch nur etwa 3 Milliarden Nucleotide, von denen nur wenige Prozent zur Codierung der zirka 50 000 Proteine des menschlichen Organismus herangezogen werden. * (Kursiv vom Verfasser)
* Dagegen ist eingewandt worden, dass wir ja mit nur wenigen binären Entscheidungen bei den Differenzierungsprozessen auskommen. Für die rund 200 verschiedenen Gewebearten des Menschen bräuchte man danach nur ein paar Dutzend Gene. Der Einwand verkennt offensichtlich die Fülle der notwendigen raumzeitlichen Makro- und Mikroordinationen auf anatomischer und biochemischer Ebene. Man denke nur einmal an die Details des menschlichen Auges. Im übrigen ändert die Frage nichts an der Tatsache der plasmatischen Genregulation.
Damit möchte Eigen zeigen, dass wir heute noch viel zu wenig wissen, um exakte Wahrscheinlichkeitsberechnungen zur Entstehung des Lebens vornehmen zu können. (Aber mit 50 000 spezifischen Proteinen hat man schon einmal eine sehr gute Basis für vernünftige Überlegungen; - vgl. Vollmerts Antwort zur Eigenschen Kritik der Wahrscheinlichkeitsansätze 1985). Richtig ist jedoch der oben zitierte Gedankengang, dass durch die genetische Information nicht alle Strukturen der Lebewesen beschrieben werden können, - womit wir wieder zu der Frage nach der zytoplasmatischen Information zurückkehren, um mit dem folgenden Zitat und der anschließenden Tabelle noch einmal an einige neuere Befunde zu erinnern.
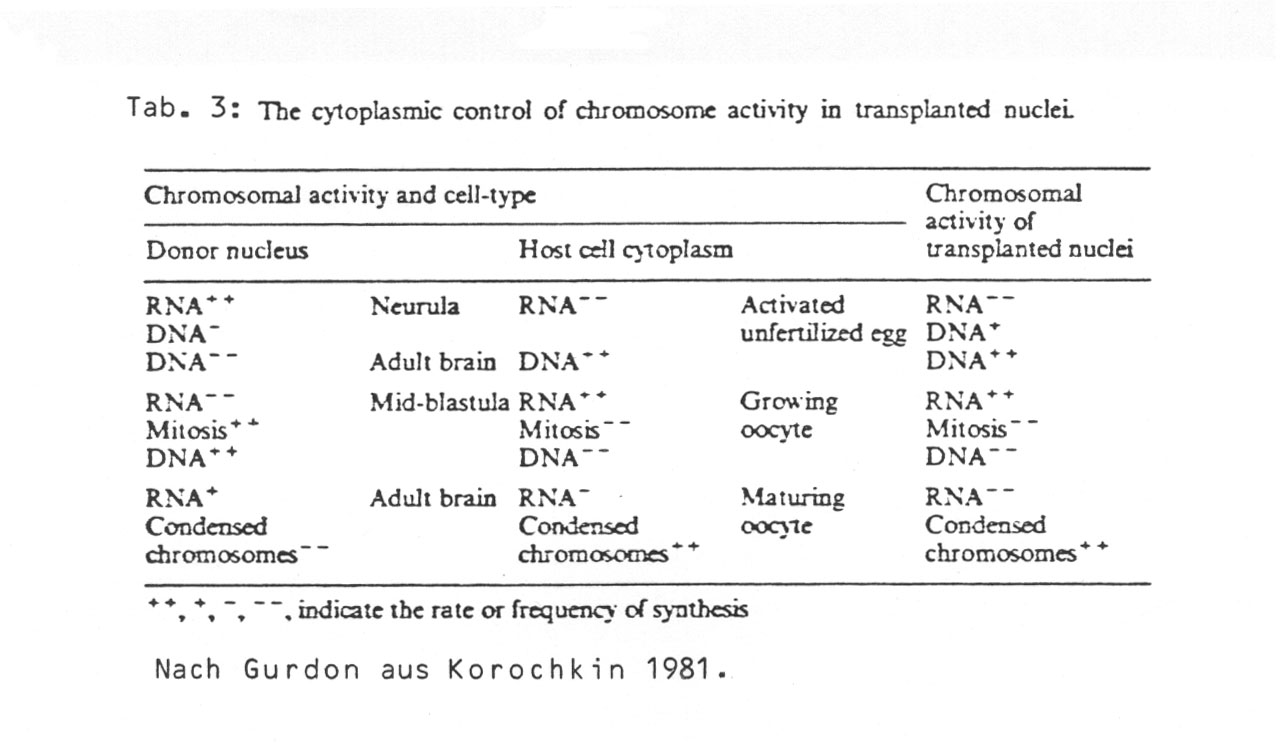
Korochkin fasst das Thema CONTROL OF NUCLEAR FUNCTION BY CYTOPLASMIC FACTORS (1981, p. 133) wie folgt zusammen:
In Gurdon's experiments (Gurdon, 1970, 1971) transplantation of amphibian nuclei convincingly demonstrated that factors in the cytoplasm regulate nuclear function. For instance, autoradiography showed that the neurula stage nucleus which synthesizes all types of RNA's will terminate this synthesis 1 h after its transplantation into egg cytoplasms. RNA and DNA synthesis rates were determined with the aid of labeled precursors (Table 3).
As seen in this table, the activity of the transplanted nucleus corresponded to the condition of the cytoplasm into which this nucleus is transplanted. The polyacrylamide gel of the RNA's synthesized by this nucleus demonstrates that this RNA synthesis is controlled by the surrounding cytoplasm.
Gurdon weist (1986, p. 306) darauf hin, dass die inzwischen feiner gewordenen molekularbiologischen Nachweismethoden die bisherigen Ergebnisse weiter fundiert haben. Die verschiedenen RNA-Klassen können überdies jetzt noch früher in der Entwicklung nachgewiesen werden. "In all experiments nuclear transplant embryos change their pattern of synthesis to coincide exactly with that of fertilized controls at the same stage." Bei der Klärung zweier scheinbarer Ausnahmen spricht der Autor von "der Regel der Zytoplasmatischen Dominanz" (p. 309).
Nagl schreibt 1986, p. 226:
It has been pointed out (Nagl 1979) that gene products cannot control differentiation and morphogenesis as basic mechanisms as their transcription requires a regulator itself, which is again a gene product and therefore requires regulators, etc., ad infinitum. Also, hormones can only "regulate" (better "induce" or "amplify") some events if the cell provides a receptor, i.e., the cell is already differentiated. These and many other aspects of developmental biology and regeneration indicate the necessity of regulatory information which is different from the genetic information. This opinion does not exclude the fact that hormones induce competence and hence the formation of further receptors, and that, in a determined or predifferentiated cell, autoregulative mechanisms are effective, as in bacteria.
Lovtrup bezieht eine ähnliche Position wie Heitler, Eigen und Nagl, wenn er den Fragenkomplex (1974, p. 400) folgendermaßen kommentiert:
...a link, currently neglected, is required to bridge the realm represented by the genetical information and that of the living organisms (Schützenberger, 1967).
(p. 402:) Probably the various extranuclear factors in the embryonic substrate exert functions which are so essential to the normal cause of epigenesis that very slight variations may have deleterious effects. This circumstance must seriously hamper the study of their inheritance.
(Ähnlich Lovtrup 1977, 1981, 1982, 1985, 1987.)
Derselbe Autor bemerkt 1984, p. 170:
...now it is claimed by many that all biological information resides in the genome. That this statement is made at all is astounding, because it is so easily refuted.(...lit.).
I will stress that the structural information "to be a cell" is contained in the fertilized egg, and is transmitted to other cells through cell division. The information required for cell division presupposed the existence of a cell, but that is not enough, further necessary information is embodied in the centriole, which usually is supplied by the fertilizing sperm. Cell and centriole are indispensible elements in the process of cell division, but they are assisted by other elements at lower levels of organization, like microtubules and actin filaments.
Und p. 172:
It is a generally accepted dogma that, barring mutations, the genes are faithfully reproduced during mitosis, and that consequently the genomes are identical in all nuclei in any individual organism. On this premise it follows that the phenomenon of cell differentiation must be governed by extragenomic, and most likely by extranuclear factors. [weitere Punkte beim Verfasser]
Welcher Natur könnte dieses Informations- und Regulationssystem sein?
Zu dieser Frage seien einige Überlegungen von Sager (1972, p. XI) zitiert (die sie jedoch selbst noch als Spekulation bezeichnet):
...the possibility remains open that further classes of cytoplasmic genes may exist, beyond those in chloroplasts and mitochondria, of DNA's associated with other cytoplasmic organelles, Structures, or membrane systems.
(p. 365:) Indeed, it is a reasonable hypothesis that additional DNA's may yet be found in the cytoplasm associated with particular membrane fractions such as the plasma membrane, Golgi apparatus, and endoplasmatic reticulum, just as DNA's have been associated with chloroplasts and mitochondria. Such DNA's could play a part in maintaining the specificity of various membranes of the eukaryotic cell by coding for special proteins, and these in turn could permit or block the insertion of other components from the surrounding cell sap during membrane growth.
Wenig war jedoch in den letzten Jahren zu dieser Frage zu hören. Nagl (1980) mahnt bei diesem Thema zur Vorsicht. Koch et al. haben 1983 zwar dazu einige Beobachtungen gemacht, sie berichten jedoch von Hinweisen, dass die von ihnen im Plasma nachgewiesene (nicht-mitochondriale) DNA aus embryonalen Zellkulturen des Huhns aus dem Zellkern stammt, sich aber in ihrer Struktur von der Kern-DNA unterscheidet und beschließen ihren Bericht (1983, pp. 253/254):
Because we cannot offer a functional aspect, we do not wish to speculate further on the possibility that cytoplasmic DNA is a possible carrier of information across the nuclear and cellular membranes and results from gene amplification during cell differentiation, etc. Nevertheless, the reported undermethylation of cytoplasmic DNA is a most remarkable feature in view of recent studies indicating that DNA methylation is a key event in control of gene function.
Trotz dieses Berichts sind die bisherigen Ergebnisse zu diesem Thema wenig befriedigend, was wohl z.T. mit unzureichenden Nachweismethoden zu tun hat.
Maclean weist 1977, pp. 109/111 darauf hin, dass Bakterien nach vollständigem Zellwandverlust durch anhaltende Penicillinbehandlung ihre Zellwand nicht mehr resynthetisieren können. "The enzymes which are responsible for laying down the murein layer apparently require an existing murein template on which to build additional material. Loss of template means permanent loss of the structure." Weiter ist bei jedweder Polysaccharidsynthese ein "primer" in Form des Produkts notwendig (bei der Glykogensynthese wenigstens 4 Zuckereinheiten). "We can surely conclude that if, during development, any cell is left without glycogen, even the activation of all nuclear genes will not achieve further glycogen synthesis." (Beide Zitate p. 111.)
Einen weiteren Ansatz findet man in gewissen autoregulatorischen Fähigkeiten der Zellmembran. Sandhoff weist (1980, p. 431) darauf hin, dass 'ein Zusammenbruch membrangebundener Glykolipide, soweit sie von membrangebundenen Enzymen katalysiert werden, von der Membran selbst reguliert wird'. - Natürlich müssen die Enzyme selbst 'irgendwo' kodiert und synthetisiert werden - womit wir wieder beim DNA-Thema sind.
Bemerkenswert ist in diesem Zusammenhang auch das nachstehend aufgeführte Beispiel der Cytoskelett-Organisation, deren Vererbung nichts mit der DNA zu tun hat (weitere Fälle bei Markert und Ursprung 1971). Die darauf zitierten Entwicklungsmöglichkeiten und maternelle Vererbung bei der Schnecke Limulaea peregra heben wieder die Bedeutung der ooplasmatischen Konstitution (diesmal) in Verbindung mit der Cytoskelettorganisation hervor. Alberts et al. (dort weitere Literatur) fassen die Fragen (1983, p. 607) wie folgt zusammen:
...the best-documented example of cytoskeletal inheritance comes from studies of Paramecium, a large single-celled organism whose surface is covered with rows of motile cilia. Normally, all of the rows are aligned in parallel, enabling a highly coordinated beating of the cilia. By experimental manipulations, it is possible to disturb this pattern and produce some inverted rows of cilia that beat in the opposite direction to that of their neighbors. Once established, such altered patterns are passed on to the cells of successive generations, apparently forever. This form of heredity has nothing to do with DNA: the modified cells inherit a particular pattern of ciliary rows via their cortical cytoskeleton.
The ability to inherit directly a particular cytoskeletal organization could have far-reaching consequences for the morphogenetic processes responsible for shaping the adult body plan of an organism. A clear example is seen in the development of the snail Limulaea peregra, whose coiled shell is genetically determined to be either a right-handed or a left-handed spiral. The handedness of the adult is observable as early as the eight-cell stage of development, in the asymmetric pattern of cell divisions known as spiral cleavage. This handedness of the embryo is determined by a slightly skewed position of the mitotic spindle during the early cleavages, which in turn is determined by maternal inheritance: the genotype of the mother snail (whether right- or left-handed) determines the form of the egg, independent of the form of her own shell. In the course of oogenesis, a factor is presumably produced by the mother snail that specifies the position of the mitotic spindle in the mature egg, and this factor determines the handedness of the embryo independent of the embryo's own genes.
Examples such as these show that the cytoskeleton of one cell can influence that of its daughter cells and affect the morphology of an entire organism. How such organizational information is propagated from cell to cell is still a mystery. (Kursiv von den Verfassern.)
Die Summe der Ergebnisse zum Thema "Regulation über das Plasma" spricht, generell formuliert, für eine überragende Rolle des Eiplasmas und des Plasmas überhaupt sowohl für die Genreplikation als auch für die Genregulation. Auf molekularbiologischer Ebene werden dafür zur Zeit stabile m-RNAs, genotrope Proteine, Kortexstrukturen und die DNA-unabhängige Vererbung von Cytoskelettelementen diskutiert, wobei für jeden dieser Punkte zahlreiche Untersuchungen und Befunde vorliegen. Darüber hinaus deuten einige Ergebnisse auf einen cytoplasmatischen (außerhalb der Chloroplasten und Mitochondrien gelegenen) DNA-Pool hin, der hier ebenfalls von Bedeutung sein könnte.
Kommen wir auf die Fragen nach dem Artbegriff und dem Ursprung der Arten zurück. Wir hatten schon hervorgehoben, dass von neodarwinistischer Seite dem Plasmon keine bedeutende Rolle (wenn überhaupt eine) zu diesen Fragen beigemessen wird und wollen das mit zwei Zitaten dokumentieren. Mayr behauptet 1967, p. 142 zu seinen Ausführungen:
Absichtlich hat diese Zusammenstellung bisher das Vorhandensein von genetischem Naterial ("Plasmagene") im Zytoplasma übergangen. Das wenige, was darüber bekannt ist, spricht nicht für eine große evolutive Rolle, so wichtig es auch für die Physiologie der Organismen sein mag.
Lüers, Sperling und Wolff sind (1974, p. 195) noch deutlicher, wenn sie schreiben:
Vor 10 Jahren wäre es noch erforderlich erschienen, die Frage einer evolutiven Bedeutung nicht-mendelnder Erbphänomene ausführlich mit Beispielen zu diskutieren. Aus dem Pflanzenreich hätten die Fälle von erblichen Plastiden- und Plasmonunterschieden behandelt werden müssen, die vom Zellkern weitgehend unabhängig sind. Die von DARLINGTON (1939) allgemein als "Plasmagene" bezeichneten plasmatischen Bestandteile mit Nukleinsäurecharakter gehören hierher. Konkreter wären als Beispiele aus dem Tierreich bei Paramecium der "killer-Faktor" und seine Mutationen sowie die "mate-killing"-Eigenschaft anzuführen, die plasmatisch, aber nicht unabhängig von Kerngenen vererbt werden. Bei Drosophila sind es die biologisch und genetisch äußerst interessanten Phänomene der CO2-Empfindlichkeit als ältestes Beispiel plasmatischer Vererbung bei einem tierischen Vielzeller (L'HERITIER und TEISSIER, 1937) und die Wirkung bestimmter "sex-ratio"-Faktoren (SR). Sie alle erfüllen zwar eindeutig die Bedingungen extrachromosomaler Vererbung, jedoch sind für das killer-Merkmal, die CO2-Empfindlichkeit und die SR-Faktoren experimentell übertragbare infektiöse Agenzien als Ursache nachgewiesen worden: bei den beiden ersten sind es Partikel von Virusnatur, beim SR-Phänomen eine Spirochaete. Im letzten Fall steht eine exogene Infektion außer Frage, bei den anderen ist sie in Betracht zu ziehen. Es ist aufschlussreich, wenn bei dieser Lage in der Monographie über die "Extrachromosomale Vererbung" von JINKS 1967 geschrieben wird: "Bisher haben in der Natur vorkommende Plasmonunterschiede im Tierreich nur bei einer Art nachgewiesen werden können, der Stechmücke Culex pipiens. Gemeint sind die subtilen und informationsreichen Analysen von Kreuzungen geographischer Rassen dieser Art durch LAVEN (1967) mit dem Ergebnis reziprok unterschiedlicher Kreuzbarkeit und einer unterschiedlichen Bedeutung der weiblichen und männlichen Gameten für das Cytoplasma der Zygote.
Früher und wohl auch heute noch waren bzw. sind viele Biologen - noch unter dem Einfluß von PLATE - überzeugt, daß die im meso- und makrophyletischen Geschehen entstandenen Erbunterschiede auf Unterschieden im Plasmon beruhen, während die intraspezifischen Differenzierungsprozesse einem kerngebundenen Mutationsprozeß zugesprochen werden. Diese Auffassung ist heute als irrig erkannt. Sie beruhte auf einer ungenügendem Kenntnis des Verhaltens reziproker Bastarde bei den entsprechenden Kreuzungsansätzen.
Trotzdem kommt den plasmatischen Unterschieden, also dem plasmatischen Mutationsprozeß im Evolutionsgeschehen in einer Hinsicht sicherlich hohe Bedeutung zu. Unverträglichkeiten zwischen verschiedenen Genotypen und Plasmatypen bei Rassen- oder Artkreuzungen sind mehr oder weniger strenge Kreuzungsbarrieren. Sie tragen zur genetischen Isolierung solcher Gruppen bei. Die Plasmonunterschiede wirken also als Isolationsbarrieren. Wahrscheinlich sind die bekannten Ergebnisse in den Rassen- bzw. Artkreuzungen mit den Schmetterlingen Lymantria (GOLDSCHMIDT, 1934) wie Solenobia (SEILER und PUCHTA, 1956) mit der Entstehung fortpflanzungsfähiger, weil intersexer Nachkommen Beispiele dafür. Für die Entstehung neuer taxonomischer Unterschiede sind jedoch plasmatische Divergenzen nach heutiger Auffassung bedeutungslos.
Zu diesen Ausführungen sei zunächst auf einen gravierenden Widerspruch hingewiesen: In der gesamten neodarwinistischen Literatur wird den Isolationsbarrieren für Artbegriff und Evolution die größte Bedeutung zugewiesen, "für die Entstehung neuer taxonomischer Unterschiede sind jedoch plasmatische Divergenzen nach heutiger Auffassung bedeutungslos." (Ebenso Jinks 1967, p. 158 )
Warum - fragt man sich - wird all den anderen, oben ausführlich diskutierten Isolationsbarrieren eine so große Bedeutung für die Entstehung neuer taxonomischer Unterschiede beigemessen, wenn gleichzeitig die durch plasmatische Faktoren verursachten Isolationsbarrieren keinerlei Sinn für das Ursprungsthema haben sollen?
Mit dem ohne Frage wichtigen Nachweis von sekundären (d.h. nicht-genetischen) plasmatischen Veränderungen (killer-Merkmal, CO2-Empfindlichkeit, SR-Faktoren) durch Viren und Spirochaeten (inzwischen auch für den Fall von durch Rickettsien, Literatur bei Beale und Knowles 1978) wird auch die Bedeutung der primären, genetisch-plasmatischen Barrieren verneint. Diese beiden Bereiche liegen jedoch auf völlig verschiedenen Ebenen. Vermutlich steht hinter dieser Ablehnung die Abneigung gegen Plates Lamarckismus und das Ziel, ähnliche Theorien von vornherein zu unterbinden. Lamprechts Lamarckschen Ansatz wollen wir unten diskutieren.
Weiter sollte man bei der von Lüers et al. gegebenen Bestandsaufnahme berücksichtigen, dass ja schon sekundäre Ursachen wie die genannten Viren und Spirochaeten entscheidende Regulator-Wirkungen über das Plasma ausüben können - wieviel mehr muss das dann innerhalb des Systems durch primäre genetisch-plasmatische Ursachen möglich sein!
Im Gegensatz zum zitierten neodarwinistischen Trend sind zahlreiche Autoren aufgrund ihrer Untersuchungen und Befunde zu einer hohen Bewertung des Cytoplasmas für den Artbegriff und den Ursprung der Arten gekommen.
Zu der folgenden Dokumentation sei vorausgeschickt, dass es bei den meisten dieser Zitate noch gar nicht um die Frage nach einem unabhängigen, übergeordneten Regulationssystem geht (wie wir das oben im Detail diskutiert haben), sondern um den bereits gut erforschten Teil der Plastiden- und Mitochondrien-Genetik.
Nachstehend sei wieder eine Serie von Zitaten (zunächst) kommentarlos aufgeführt. Die Hauptpunkte sollen anschließend diskutiert werden.
Kheyr-Pour (1986, p. 259):
It is now realized that the cytoplasm can affect disease reaction, e.g. in corn, and has a role in the evolution of higher plants.
Kihara (1982, p. 449):
There must be a close relationship between cytoplasm and nucleus, in which even a small change (mutation) in the nucleus has to be compatible with cytoplasm for adaptation.
...the evolution of plant species must be the result of coordination of hereditary units, genome and plasmon.
Grun (1976, p. 347) :
Both the nuclear genotype and the cytoplasmic factors are capable of change, and there is no way of knowing now which was the initiator of change and which the respondent. A given change in mitochondrial or plastid genotype could have become established only if the nuclear genotype was preadapted to it or if a nuclear adaptation followed rapidly.
Irrespective of which changes first, the interaction results by its nature in a more conservative evolutionary process than any component might undergo if it evolved independently. Some types of nuclear or cytoplasmic mutants that might otherwise have been evolutionary successes have probably not been established because they could not collaborate well with the population of other factors present in the cell at the time in question. The response of one part of the total cellular genotype to change in other parts may well be one of the limitations to the evolution of the organism.
In der Einleitung (pp. V/VI) schreibt derselbe Autor nach Hinweis, dass sich die Evolutionslehre bislang hauptsächlich mit den Wechselwirkungen zwischen den Kerngenen und Fragen zum Verhältnis Organismus und Umwelt beschäftigt hat, über den Zweck seines Buches:
The focus is on the fact that the fitness of the organism to survive here is determined by the interactions among its diverse genetic centers, all of which are acted upon during selection and none of which can respond independently of the others.
Thus, if a chromosomal mutant is produced that conditions better disease resistance in a plant, natural selection might ordinarily favor that mutant in the population so that it would become the "wild" type. It must pass a test first, however: The enzyme produced by it must not interact unfavorably with the enzymes encoded by the mitochondrial genes * or the plastid genes, for if it does the mutant plant will not survive. A new chromosomal mutant that might otherwise be favorable to the survival of a mouse will be established in a wild population only if it interacts well with mitochondrial gene products and if it does not activate the leukemia or the mammary tumor viruses that are latent parts of the mouse's normal inheritance.
Conversely, a new mutant form of mitochondrion that can divide more rapidly than its brethren might show up in a plant cell and could become the established sort. If the rapid reproduction occurs at the expense of poor interaction with the chromosomal gene products or plastid gene products, the mutant mitochondrion might well destroy the cell and so never survive in the plant. If it survived, however, its effect might be that the plant would not reproduce and hence the mutant mitochondrion would be lost to the population. The evolution of each organelle, therefore, puts constraints on the evolution of each other organelle even as the changes of the environment affect them all.
*Beale und Knowles (1978, p. 41): The two genetic systems - nuclear and mitochondrial - interact intimately at several levels: firstly, the level of the mitochondrial DNA, whole structure is controlled by itself, but whose activity is controlled by products of the nuclear system; secondly, the level of the mitochondrial ribosomes, part of whose structure is controlled by each system and whose activity also by both; and thirdly, the level of the inner membranes, which contain an integrated patchwork of products of the nuclear and mitochondrial genetic systems. All this must require an extraordinary degree of mutual adaptation and correlated evolution. Our knowledge of the actual machinery of the mitochondrial genetic system is still extremely slight.
Schwanitz (1974, p. 114) [im selben Werk wie Lüers, Sperling und Wolff (1974)]:
Es steht seit Jahrzehnten fest, daß der Zellkern keineswegs der einzige Träger genetischer Information ist, sondern daß auch im Plasma Faktoren vorhanden sind, die einen beträchtlichen Einfluß auf die Vererbungsvorgänge und damit auf die Evolution haben.
(pp. 122/123:) Es ist erwiesen, daß von extrachromosomalen genetischen Faktoren wichtige Eigenschaften der Pflanze, wie die Gynodiözie, das Wachstum, die Wuchsform, die Verzweigung, die Blütenbildung, die Geschlechtsform, der Photoperiodismus, die Vitalität sowie zahlreiche physiologische Eigenschaften mitbestimmt werden können (Hagemann, 1964). Sie müssen daher für die Evolution größte Bedeutung haben können, und so ist die Frage nach dem genetischen Verhalten recht wesentlich.
(p. 127:) Schon die Untersuchungen von F.v.Wettstein haben gezeigt, daß Genotypus und Plasmotypus bei der Herausbildung der Merkmale eng zusammenwirken. Eine Disharmonie zwischen diesen beiden Elementen der Vererbung kann sich im Phänotypus des betreffenden Individuums auswirken und zu Störungen im physiologischen Verhalten führen.
...[Es] hat sich verschiedentlich gezeigt, daß die Einlagerung eines Genoms in ein fremdes Plasmon bei bestimmten Genen eine Steigerung der Mutationsrate mit sich bringen kann (Kihara, 1951; Michaelis, 1953).
Mather (1965, p. 45):
If differences between more widely separated groups depend more often on differences in cytoplasmic determinants than those between less widely separated groups, we must recognize a process of macro-evolution which differs from micro-evolution in depending characteristically on cytoplasmic as well as genic changes. It could not therefore be a simple extension of the evolutionary changes which have been studied and shown to depend essentially on nuclear adjustments.
Mather fügt auf derselben Seite folgenden Rat hinzu:
But to be useful as a hypothesis, it must be open to test by accumulation of appropiate evidence. The danger would he in invoking the supposed novelty always at a point beyond which information and experience will carry us, so that it would be permanently immune from test. It would then cease to be a helpful hypothesis and indeed has only to be stated in this form to be discredited.
Dieser Rat trifft grundsätzlich auf jede Hypothese zu, zumal auf den neodarwinistischen Ansatz mit seiner Extrapolation der mikroevolutionistischen Ergebnisse auf den Ursprung aller Lebensformen und seinem ganzen Satz von Immunisierungserklärungen, bei der die Nicht-Reproduzierbarkeit seiner Aussagen für die Entstehung höherer systematischer Gruppen integraler Bestandteil durch die postutierten Zeiträume ist.
Zur Frage nach Kernveränderungen im Zusammenhang mit der Differenzierung schreibt derselbe Verfasser (p. 54):
...however important nuclear changes may be in the process of differentiation, they themselves must always be the consequences of some other antecedent cause. The cause of their occurrence would thus seem to lie outside the chromosomes, in fact in the cytoplasm to which we must turn now.
Wie wir oben schon ausführlich dokumentiert haben, sprechen die gründlichen Untersuchungen vieler Forscher an den unterschiedlichsten Organismen für die Richtigkeit dieses Ansatzes. Neben den bereits gegebenen Literaturangaben können zahlreiche weitere Untersuchungen für Kern-Plasma-Interaktionen zitiert werden (z.B. Beletzky 1978, Labarene und Bernet 1979, Ahokas 1980, Dujardin et al. 1980, Fukunaga 1980, Hachtel 1980, Rao und Fleming 1980, Sears 1980, Hinton 1981, Mariol 1981, Del Guidice 1981, Thiellement 1982, Lüning 1982, 1983, Browder 1984, Dean et al. 1985, Edgar et al. 1986, Lorberboum et al. 1986, und viele weitere Arbeiten).
Bei genauer Betrachtung der zuletzt aufgeführten Serie von Zitaten zur Bedeutung des Plasmons für den Artbegriff und den Ursprung der Arten zeigen sich für die Synthetische Evolutionstheorie mehrere Gefahrenquellen, die wir im folgenden kurz erläutern möchten.
Grun hat in seiner Arbeit nachdrücklich auf die wechselseitige Abstimmung der verschiedenen Genzentren der Zelle und auf die dadurch bedingte Limitierung genetischer Veränderungen hingewiesen und hat diesen Punkt detailliert begründet. "...the interaction results by its nature in a more conservative evolutionary process than any component might undergo if it evolved independently." - Es ist klar, dass eine solche (wenn auch nur relative) Limitierung beim Versuch, den Ursprung aller Lebensformen mit den uns bekannten definitionsgemäß richtungslosen (Kern-)Mutationen und Selektion zu erklären, wenig hilfreich ist. Gibt es doch schon mehr als genug Schwierigkeiten, die Ursprungsfrage ohne eine solche Limitierung zu erklären (vgl. z.B. Dewar 1957, Lönnig 1976, Illies 1983, Kahle 1984, Arthur 1984, Ho und Saunders (Eds.) 1984, Reid 1985).
Wir möchten jedoch nachdrücklich hervorheben, dass diese Limitierung durch eine Fülle von Untersuchungen und Ergebnissen wissenschaftlich so gründlich abgesichert ist, dass hier kein vernünftiger Zweifel mehr erhoben werden kann. Durch die von Grun und anderen Autoren beschriebene Limitierung ist allerdings den Organismen eine größere Stabilität zugesichert als manchem Evolutionstheoretiker nach seinen bisherigen Voraussetzungen und Zielen wünschenswert erscheinen dürfte.
Mit dem oben postulierten (dem Kern übergeordneten) meiotisch-plasmatischen Kontrollsystem eröffnen sich gleich mehrere weitere Probleme für den Neodarwinismus. Einmal deuten die bisherigen Befunde auf eine Steigerung der 'Art'-Stabilität hin, die noch weit über die Grunsche Limitierung hinausgeht. Diese Konstanz könnte im grundsätzlichen Widerspruch zum neodarwinistischen Ansatz stehen, dafür jedoch das die gesamte Paläontologie durchziehende Phänomen der Stasis (vgl. z.B. Harlandt et al. 1967, Eldredge und Gould 1972, Stanley 1979, Eldredge und Stanley (Eds.) 1984) erklären. Auch dürfte eine solche Konstanz dem Grundgedanken aller schöpfungstheoretischen Überlegungen entsprechen.
Mather weist auf den durch ein autonomes plasmatisches System gegebenen grundsätzlichen Unterschied zwischen Mikro- und Makroevolution hin (wie vor ihm Lamprecht und Plate). Ein Hauptsatz der Synthetischen Evolutionstheorie ist jedoch die Erklärung aller Lebensformen durch Extrapolation der mikroevolutionistischen Ergebnisse.
Eine Fülle von Tatsachen weist jedoch darauf hin, dass mit diesem Problemkreis noch zahlreiche offene Fragen verbunden sind, die man (durch die oben zitierte Verneinung des Problems) längst für erledigt hielt, um mit Mutation und Selektion eine im Prinzip vollständige Antwort auf die Ursprungsfrage geben zu können (ich möchte auf diesen Punkt später noch einmal detailliert zu sprechen kommen).
Ärgerlich dürfte für die meisten Biologen der erneute Versuch sein, auf der Plasmonbasis eine Art Neolamarckismus zu begründen. Trotz aller gegenteiligen Behauptungen ist jedoch auch dieses Thema bis heute nicht zur Ruhe gekommen (vgl. Steele 1980, Grassé 1985).
Lamprecht beantwortet die Frage nach der Entstehung der Arten folgendermaßen (1966, p. 425):
Hierfür verbleibt nur die Annahme, daß die Arten von der Umwelt Eindrücke, Impulse erhalten, die sie registrieren und über anscheinend beliebig lange Zeiträume magazinieren können. Und diese Registrierung von Umweltimpulsen kann selbstverständlich nur stofflich erfolgen. Sie muß auch von Generation zu Generation weitergegeben werden können. Man könnte dies als das materielle Gedächtnis der Organismen bezeichnen. Und wo diese Impulse regestriert und magaziniert werden können, diesbezüglich gestatten die mitgeteilten Kreuzungsergebnisse nur eine Möglichkeit: Es können weder die Gene, noch die Progene in Frage kommen, sondern nur das Plasma.
Stellt man dazu konkret die Frage, wie mit einem solchen Mechanismus etwa der Fangapparat von Utricularia vulgaris und anderer tierfangender Pflanzen enstehen konnte (oder überhaupt die Synorganisation im Tier- und Pflanzenreich), so dürfte klar sein, dass man mit dieser Erklärung nicht weiterkommt.
Wir möchten hier noch einmal feststellen, dass wir die inzwischen wiederholt reproduzierten empirischen Befunde Lamprechts zum Themenkreis der interspezifischen Gene und des meiotisch-plasmatischen Kontrollsystens scharf von einigen fragwürdigen Hypothesen wie den ausschließlich auf die "Progene" beschränkten Mutationen und seinem lamarckistischen Ansatz zur Entstehung der Arten trennen müssen, wenn wir naturwissenschaftlich gesicherte Ergebnisse nicht mit Interpretationsfehlern über Bord werfen wollen.
Nach allen bisher vorliegenden Untersuchungen sind unüberbrückbare Artbarrieren tatsächlich durch artentrennende Gene und Gensysteme in Korrelation mit dem meiotisch-plasmatischen Kontrollsystem gegeben: Durch 'interspezifische' (Regulator)-Gene und Gensysteme bedingte Merkmale bzw. Merkmalskomplexe können nicht zusammen mit Fertilität von der einen Art in die andere (und vice versa) überführt werden.
Als weitere im Gegensatz zur Synthetischen Evolutionstheorie stehende Auffassung sei noch der Neovitalismus genannt, der von einigen Autoren mit der Eigenständigkeit des Cytoplasmas verbunden wird (Driesch 1894, 1929, Heitler 1961/1970, 1976, Trincher 1981 und andere). Auf diese Frage möchte ich jedoch an dieser Stelle nicht weiter eingehen. [Ein umfangreiches Thema für sich, das sowohl fragwürdige als auch eindeutig positive Elemente enthält.]
Halten wir fest, dass mit der Eigenständigkeit des Cytoplasmas sich Möglichkeiten der Theorienbildung eröffnet bzw. wiedereröffnet haben, die man von Seiten der Synthetischen Evolutionstheorie längst überwunden glaubte und die darüber hinaus den ganzen neodarwinistischen Ansatz in Frage stellen können. Hier dürfte die Hauptursache für die Abwertung plasmatischer Strukturen für den Artbegriff und den Ursprung der Arten bei vielen Befürwortern dieser Evolutionstheorie liegen.
Im Gegensatz zum Neodarwinismus haben sich Forscher wie Barry immer wieder wie folgt zum 'Kernmonopol' geäußert (1986, p. 317):
...the implication that DNA is an almost unique repository of 'information' within the cell appears to arise either from ignorance of cell function or from a misunderstanding of information theory according to which 'information' can be contained not only in words and other linear combinations of alternative symbols, but in any non-random structure. Hence, in a living cell 'information' can be, and is, also contained in other cell molecules and structures derived from them.
Werfen wir zum Schluss dieses Kapitels noch einen Blick auf eines der vielen noch ungelösten Probleme der Meiosis.
Alberts et al. stellen zur Funktion der in der Meiose auftretenden Lampenbürstenchromosomen 1983, p. 454 fest:
In some respects, the pattern of transcriptional activity on lambrush chromosomes does not seem to be typical of an ordinary somatic cell. First, all the loops are usually heavily transcribed, and many produce unusually long primary transcripts. Second, highly repeated DNA sequences that are thought not to be transcribed in normal cells are transcribed from some lambrush loops (Figure 8 - 103). Finally, at least in Xenopus, most of the enormous transcriptional activity of the lambrush chromosomes seems not to lead to mRNA production, since all of the stored mRNA molecules for the egg have already been made by the time the lambrush chromosomes reach their largest and most active phase. Thus, the function of much of this activity is mysterious and may be unique to the oocyte.
Durch die neueren Forschungen ist das Bild zur Genregulation in der Ontogenese und Meiose sowie des Aufbaus spezifischer Strukturen und Prozesse während der Meiose selbst immer komplexer geworden (vgl. zum letzteren Thema z.B. Bouchard und Stern 1980, Rasmussen und Holm 1980, Carpenter 1981, Cheng und Bassett 1981, Goldstein und Trianthophyllon 1981, Kumar et al. 1981, Laurie and Jones 1981, Zickler und Sage 1981, Lie und Lane 1982, Moses 1982, Dietrich und Mulder 1983, Cardoso et al. 1984, Hotta et al. 1984 und viele andere).
Diese Arbeiten beschäftigen sich meist noch mit so bekannten Fragen, wie der Bildung des synaptischen Komplexes, der Reduktion und Rekombination. Aber selbst hier liegen noch zahlreiche offene Probleme im Ablauf der biochemischen Prozesse vor. Die Erforschung der spezifischen Unterschiede zwischen Oo- und Spermatozytenbildung hält - wie mit dem obigen Zitat angedeutet - noch viele offene Fragen bereit, insbesondere das Problem der langwierigen Syntheseprozesse während der Oogenese.