
(6) STERILITÄT DURCH CHROMOSOMENMUTATIONEN (PUNKT 3, p. 68)
"Die Hybriden sind steril, weil sich ganze Chromosomen, Chromosomensegmente oder Genkomplexe ungleichmäßig auf die Gameten verteilen" (Stebbins, vgl. p. 68 dieser Arbeit). Grant gibt (1976, 1981) zu diesem Thema einen guten Überblick:
Abb. 25: Typen von Chromosomenumbauten. Das strukturell geänderte Chromosom (Bezeichn. Rear.) wird jeweils der entsprechenden Standardanordung gegenübergestellt (Std.). Die Pfeile kennzeichnen die Bruchstellen auf den Standardchromosomen, aus denen dann die jeweiligen Umbauten entstehen können. Das Zentrometer ist als mediane oder submediane Ellipse eingezeichnet. Aus Grant 1964/1976/1981.
Ein Bruch nahe dem Ende eines Chromosoms kann zu einer terminalen Defizienz führen. Zwei Brüche in einem Chromosom vermögen eine interstitielle Deletion, eine Duplikation, eine parazentrische oder eine perizentrische Inversion zur Folge haben. Drei Brüche in einem Chromosom können eine Transposition hervorruren. Wenn sich die Bruchenden von zwei nicht homologen Chromosomen vereinigen, ergeben sich kleine reziproke Translokationen oder Translokationen ganzer Chromosomenschenkel. Die Vereinigung der Bruchenden von drei oder mehr nicht homologen Chromosomen endlich führt zu aufeinanderfolgenden Translokationen (Abb. 25).
Werden durch Chromosomenmutationen neue Arten geschaffen?
Es ist keine Frage, dass durch Chromosomenmutationen die Fertilität zwischen verschiedenen Linien herabgesetzt sein kann. Wie ich vorhin schon mit Dobzhansky erwähnte, ist chromosomale Sterilität bei Pflanzen häufiger als bei Tieren anzutreffen.
Ganz energisch hat der Pflanzengenetiker Lamprecht (1966, 1974) die Frage nach dem Ursprung neuer Arten durch Chromosomenmutationen verneint. Nach jahrzehntelangen Studien der Chromosomenverhältnisse bei Pisum und anderen Pflanzenarten schreibt er (1966, p. 285) zur Frage, ob man nicht zumindest bei einer chromosomal bedingten vollkommenen Sterilitätsbarriere zweier Linien von verschiedenen Spezies sprechen könne:
Diese Frage will ich unbedingt mit Nein beantworten.
- Worauf er die Antwort an einem Pisum-Beispiel erläutert:
Um diese Verneinung zu begründen, braucht man nur an die Ergebnisse verschiedener Kreuzungen mit oect. fulvum zu denken. Sobald als zweiter Elter eine Linie gewählt wird, die in gewissen Chromosomen mit fulvum strukturell übereinstimmt, ist die Barriere gesprengt. Es resultiert unmittelbar ein Grad von geringerer Sterilität, z. B. 25%, und damit ist der Isolationsmechanismus eliminiert. Und ganz Gleiches muß natürlich für alle Fälle Gültigkeit besitzen, wo auf dem Kreuzungswege hergestellte Strukturtypen vollkommen sterile F1-Individuen geben. Es besteht hierbei eine vollkommene Analogie mit den fulvum-Kreuzungen.
Es ist hier also wieder eine gewisse Reversibilität der Verhältnisse, die mit zur Verneinung der oben gestellten Frage führt. Durch die Lamprechtschen Studien über chromosomale Reparaturen, die zu einer Erhöhung der Fertilität zwischen verschiedenen Linien geführt haben, wird diese Schlussfolgerung weiter bestärkt.
Im Gegensatz zu Lamprechts Auffassung (und der auf p. l7 zitierten Genetiker) haben neuerdings Selim und Hussein (1983) wieder versucht, dem Ökotypus P. abyssinicum Artstatus zuzuschreiben. Begründung (p. 412):
Abyssinicum is specifically distant from all other taxa. It showed striking morphological differences from other taxa, and crossability succeeded only with difficulty with Pisum sativum, var. Little Marvel.
Ich habe selbst jahrelang mit beiden Pisum-"Arten" gearbeitet (vgl. S. 17). Von besonders auffallenden morphologischen Unterschieden zwischen beiden Ökotypen kann kaum die Rede sein. Morphologische Unterschiede zwischen verschiedenen Linien innerhalb von Pisum sativum können wesentlich ausgeprägter sein als zwischen manchen P. sativum- und P. abyssinicum-Linien (man vergleiche dazu nur einmal die Blattmutanten und Blattrekombinanten sowie die Größenunterschiede innerhalb von P. sativum; - Gottschalk 1964, 1984).
So bleiben dann vor allem die die Fertilität herabsetzenden chromosomalen Unterschiede zwischen beiden Ökotypen, die nach den Worten der oben zitierten Autoren, "eine beträchtliche reproduktive Isolation anzeigen".
Abgesehen davon, dass eine beträchtliche reproduktive Isolation noch nichts über den Artbegriff auszusagen braucht - wie wir oben ausführlich für prä- und postzygotische Isolationsmechanismen gezeigt haben - übersehen die Autoren völlig, dass man einen auf chromosomalen Unterschieden beruhenden Artbegriff auch innerhalb der verschiedenen Pisum-"Arten" konsequenterweise anwenden müsste, was dann zu zahlreichen neuen "Arten" führen würde.
So geben zum Beispiel Gottschalk und Wolff (1983, p. 4) für ihre röntgen- und neutroneninduzierten Mutanten an, dass u.a. etwa 120 (!) Translokationen induziert wurden. Weiter bemerken sie:
A large number of them was maintained by propagating the translocation-heterozygous plants. In their offspring, plants homozygous for the translocated chromosomes were selected and developed into strains. Twenty-five different strains of this group were studied with regard to vitality and fertility. Only one of them was distinctly inferior to the mother variety. All the other ones were fully fertile and vital (Gottschalk 1978). This is in contrast to the situation in Drosophila where translocation homozygosity leads often to lethality.
Aufgrund der chromosomal bedingten reduzierten Fertilität zwischen diesen Linien und der Ausgangsform, könnte man nun glatt wenigstens 25 neue Pisum-Arten aufstellen, von denen einige den Sterilitätsgrad zwischen P. sativum und P. abyssinicum noch übertreffen. Gottschalks Translokationslinie 405 B beispielsweise weist in der F1 bei Kreuzung mit der Ausgangslinie nur noch 22% Fertilität auf, Lamprechts Kreuzung seiner abyssinicum-Linie 808 mit der "englischen grazilen Kneifelerbse Chelsea" (mit normaler Chromosomenstruktur) zeigte hingegen in der F1 noch 39,9 +- 2,34 % Fertilität (Details zu Kreuzungen mit Linien unterschiedlicher Chromosomenstruktur vgl. Lamprecht 1966, pp. 185-198 und 1974, pp. 577-582.)
Würde man allein bei Pisum alle durch Chromosomenmutationen gekennzeichneten Linien, die herabgesetzte Fertilität bei Kreuzung mit ihren Ausgangs- oder anderen Formen zeigen, Artstatus zuschreiben, dann würde das zu Hunderten von neuen Pisum-Arten und zu einem nomenklatorischen Chaos führen. Kein Genetiker hat jemals einen solchen Vorschlag gemacht, denn die Konsequenzen für das gesamte Pflanzenreich wären unüberschaubar.
Hervorzuheben ist für viele Translokationslinien noch die Tatsache, dass sie sich von ihren Ausgangslinien morphologisch kaum oder gar nicht unterscheiden. Bei Anwendung des chromosomalen Artbegriffs müssten damit auch Dutzende von neuen Zwillingsarten benannt werden, wobei der Artbegriff in zahlreichen Fällen wieder mitten durch morphologisch und ökologisch klar definierte Subspezies und Varietäten ginge.
Die einzige vernünftige Alternative ist, die verschiedenen Chromosomenmutationen und die damit verbundenen Sterilitätsgrade nicht zum Maßstab für den Artbegriff zu machen, sondern sie unter den Artbegriff zu subsumieren.
Das wird heute zum Teil schon praktiziert. So spricht Brooker (1982) von "Robertsonian Translocations in Mus musculus from N. E. Scotland and Orkney".
- Man beachte, dass der Autor den von den Translokationen betroffenen Populationen nicht eigenen Artstatus zuschreibt (obwohl er - ganz im Geiste der Zeit - meint, dass die stark reduzierte Fertilität bei den Hybriden eine für Artbildung notwendige, reproduktive Barriere bilden könnte. Er fügt jedoch einschränkend hinzu, dass die Ausbreitung dieses Mäusetyps durch eine solche Barriere sehr erschwert wäre).
Ebensowenig nehmen Nombella et al. (1982) aufgrund ihrer Karyotypstudien an der Haselmaus Eliomys quercinus L. (2 n = 50 bei der französischen Population und 2 n = 48 bei der spanischen durch Robertsonsche Translokation) eine Trennung in zwei verschiedene Arten vor.
Adolph und Klein berichten 1983, pp. 117/118 zusammenfassend über die Hausmaus:
Until 1969, geneticists and cytogeneticists believed that the karyotype of virtually all house mice (Mus muscu1us or Mus domesticus) consisted of 40 acrocentric chromosomes (Cox, 1926). Although animals with fewer than 40 chromosomes had now and then been found among laboratory mice (Evans, Lyon & Daglish, 1964; Leonard & DeKnudt, 1967; White & Tijo, 1967; Baranov & Dyban, 1971), they were regarded as rare exceptions. Even after Gropp, Tettenborn & Lehman (1969) discovered the first example of Robertsonian chromosomal variation among wild mice in the Valle di Poschiavo in the Alps (the mice had 14 metacentric chromosomes with each metacentric derived by Robertsonian fusion from two acrocentrics), there was no reason to believe that the variation was more than a local curiosity. Later, however, Robertsonian variation of mouse chromosomes was found not only in other valleys of the Alps (Gropp et al. 1972) but also in other parts of the world: in various regions of Italy (Capanna et al. 1976; Capanna, Civitelli & Cristaldi, 1977), Sicily (Lehman & Radbruch, 1977), the Eolian Islands and Dalmatia (Gropp & Winking, 1981), Greece (Winking, Gropp & Bulfield, 1981), southern Germany, Spain, Scotland (Adolph & Klein, 1981), India (Chakrabarti & Chakrabarti, 1977) and an island of the Antarctic region (Robinson, 1978). These findings suggested that the Robertsonian variation among wild mice represented a widespread chromosomal polymorphism similar to that found in other rodents, for example, gerbils (Wahrman & Gourevitz, 1973), Acomys (Wahrman & Gotein, 1972) and mice of the Leggada group (Jotterand, 1972).
Auch diese Autoren schlagen vernünftigerweise keine neuen Artnamen für die durch Robertsonsche Translokationen gekennzeichneten Populationen vor.
Said et al. berichten 1986, pp. 151-156 von tunesischen Hausmäusen mit 9 Paaren Robertsonscher Translokationen. "Five of the nine translocations are not known to occur elsewhere. In two populations only 2n = 22 mice were present, whereas in a third one a mixture of karyotypes (2n = 22, 38, 39, 40) was found" (p. 151). Eine Untersuchung der Proteinpolymorphismen zeigte keine Unterschiede zu den normalen Hausmauspopulationen. Die Autoren betonen, dass alle Populationen zu Mus musculus domesticus gehören.
Zur Tabakmaus, Mus poschiavinus, einer "extremen Varianten der Hausmaus" (Gropp und Winking), die aufgrund morphologischer Besonderheiten 1869 von Fatio ihren Artnamen bekam, kommentieren Gropp und Winking jedoch (1972, p. 266):
His (Fatio's) claim that this mouse should be considered as a separate species, M. poschiavinus, seems to be substantiated by the observation of a reduced fertility of the hybrids of the tobacco mouse and the laboratory mouse in consequence of the chromosomal rearrangements (literature).
Wenn das die Legitimation für den Artstatus ist, dann müssten auch alle anderen von Adolph und Klein oben aufgeführten Populationen eigene Artnamen bekommen - eine Konsequenz, der sich meines Wissens bisher niemand angeschlossen hat und die wohl kaum Aussicht auf Erfolg haben dürfte. Entweder bekommen alle Populationen mit Robertsonschen Translokationen Artstatus, oder der Tabakmaus ist dieser Status wieder abzuerkennen und sie wird als Subspezies im Sinne der tertiären Nomenklatur geführt: Mus musculus poschiavinus. Aufgrund der oben aufgeführten Tatsachen und Konsequenzen erscheint mir das letztere als überzeugende Lösung.
Im übrigen kam Ohno schon 1970 im Prinzip zum gleichen Ergebnis, als er schrieb (p. 43):
... at a number of coat color as well as enzyme loci, we found M. poschiavinus to possess known alleles of M. musculus. A series of remarkable chromosomal changes apparently have occurred in a rather short span of time, so that genetically the tobacco mouse is still a member of the ordinary house mouse species.
Weitere Beispiele:
Belcheva und Bisserkov erwähnen (1984, pp. 573-581) im Rahmen ihrer karyologischen Studien an der Hausratte in Bulgarien, dass diese Art je nach Chromosomenzahl und Karyotyp in vier geographischen Varianten vorkommt: die asiatische (2n = 42), ceylonesische (2n = 40), ozeanische (2n = 38) und die auf Mauritius vorkommende Variante (2n = 42). Eine Trennung in vier Arten wird nicht vorgenommen; es handelt sich in allen Fällen um Rattus rattus L..
Bocquet zeigt (1984, pp. 274/275) die Schwierigkeiten eines chromosomal definierten Artbegriffs auf und gibt u. a. folgendes Beispiel (p. 275):
... chez l'Isopode, asellote Jaera syei, les femelles montrent, a la première métaphase méiotique, en plus d'un trivalent sexuel toujour présent, treize bivalents sur les côtes d'Allemagne du Nord, douze bivalents en Hollande, onze bivalents entre le Boulonnais et le Cotentin, dix bivalents en Bretagne, neuf bivalents sur les côtes atlantiques entre Nantes et Royan et huit bivalents dans la region de Biarritz il ne s'agit pourtant là que de formes chromosomiques d'une seule et même espece.
Trotz der unterschiedlichen Chromosomenzahl handelt es sich hier nur um eine Art. Bei Drosophila mulleri, D. aldrichi und D. wheeleri und verschiedenen Drosophila-Arten Hawaiis ist hingegen nicht nur die gleiche Chromosomenzahl festgestellt worden, sondern auch das gleiche Bandenmuster (Bocquet); - hier werden nun die Arten nach morphologischen und anderen Kriterien unterschieden.
Wohin der morphologische Artbegriff im Pflanzenreich führen kann, haben wir u.a. am Beispiel der Habichtskräuter oben gesehen. Ein weiteres bemerkenswertes Beispiel aus der Zoologie liefern uns Colombera und Rasotto mit der Artabgrenzung beim Knochenfisch Gobius paganellus. Die Autoren schreiben u.a. (1982, p. 257):
The great number of species (over 600) so far described for the genus Gobius and the fact that Gobiidae display very similar morphological patterns together with a high intraspecific variability often make it very difficult to recognize these species on the base of sole morphological parameters (Fage 1925).
In particular Gobius niger jozo can be easily mistaken for G. paganellus.
... It seems that at a karyological level these species can be better differentiated since diploid chromosome numbers in Gobiidae range from 44 to 50 (..lit...) with 2n = 48 for G. niger jozo and 2n = 50 for G. paganellus.
Nun haben wir ja schon einige Beispiele für die Variabilität der Chromosomenzahl innerhalb von Arten aufgeführt. Was machen die Autoren, wenn sich im Zuge der weiteren Forschung bei diesem oder ähnlich gelagerten Fällen auch innerhalb der morphologisch sowieso kaum zu unterscheidenden 'Arten' zusätzlich noch unterschiedliche Chromosomenzahlen finden? Die Variabilität der Chromosomenzahlen von 44 bis 50 legt ja den Gedanken nahe, dass die Unterschiede nicht immer mit den morphologischen Artabgrenzungen kongruent sind.
Schmidt erwähnt (1985, p. 312) den Nager Gerbillus pyramidum, "der eine so stark schwankende Chromosomenzahl aufweist, daß sie von 40 in Algerien bis 52 in der Küstenebene von Israel und in der Negev-Wüste sogar bis 66 variiert" und fügt hinzu: "Dennoch handelt es sich um Tiere der gleichen Art."
Auf der Seite 36 zitiert er "die erst seit relativ kurzer Zeit nach Afrika eingewanderten Zwergmäuse ..., die nach MATTHEY (1967) eine extreme Chromosomenpolymorphie mit Schwankungen der Chromosomenzahl - mit Zwischenstufen - von 18 bis 36 aufweisen." (Sie werden hier anderen Tiergruppen mit ziemlich konstanten Chromosomenverhältnissen gegenübergestellt.) Auch diese Variabilität kommt innerhalb einer Art vor.
Frykman und Bengtsson berichten (1984) über genetische Differenzierung bei der Spitzmaus Sorex araneus (p. 259):
Two karyotypic races of the common shrew, Sorex araneus L., come into contact in the north of Sweden. The two races, called the northern (N) and the central/middle (M) karyotypic races, are chromosomally different but morphologically (SULKAVA et al. 1983) and genetically very similar. Out of 35 enzyme loci electrophoretically analyzed, only one showed a clear difference in allele frequencies between populations belonging to different races (FRYKMAN et al. 1983; FRYKMAN, in preparation).
... The chromosomal difference between the karyotypic races is of a complex Robertsonian type, with the same chromosome arms occurring in different combinations in the two races.
Nach Beschreibung der Translokation weisen die Verfasser auf den hochgradigen Polymorphismus innerhalb der Rassen hin und weiter auf eine dritte karyotypische Rasse (S) im Süden Schwedens. Zur Wirkung der chromosomalen Unterschiede auf die Fertilität bemerken sie (p. 267):
The complex chromosomal configurations in the meiosis of the hybrids will according to cytological theory, give these animals reduced fertility (or no fertility at all) and, therefore, hinder the free exchange of genetic material between the races.
Die Autoren meinen zwar mit White (1978), dass Rassen, die auf diese Weise voneinander getrennt sind, auf dem Wege zu zwei verschiedenen Spezies sein könnten, erheben sie jedoch nicht aufgrund der bekannten chromosomalen Differenzen in den Rang eigener Arten. Mit den Beispielen der geographischen Isolation (p. 112f) sowie der verschiedenen bisher diskutierten prä- und postzygotischen Isolationsmechanismen sei nochmals an die Fragwürdigkeit dieses Ansatzes zur Artentstehung erinnert. Im Übrigen sei noch erwähnt, dass Fryman und Bengtsson aufgrund karyologischer und Allelfrequenzuntersuchungen Genfluss zwischen den Rassen festgestellt haben (Hybridzone).
Herabgesetzte Fertilitat zwischen Chromosomenrassen gibt es auch beim blauen Polarfuchs Alopex lagopus (Canis lagopus). Christensen und Pedersen berichten (1982) über die Variation in der Chromosomenzahl beim Blaufuchs und ihre Wirkung auf die Fertilität (1982, p. 211):
Investigations in Denmark, Sweden, Finland and Poland (lit.) have shown that the chromosome number varies in the blue fox. There occur animals with 48, 49 or 50 chromosomes. Those with 50 chromosomes have 4 acrocentric chromosomes, those with 49 have 2, and finally those with 48 have no acrocentric chromosomes.
(p. 214:) The chromosome polymorphism consists of the centric fusion of the only two acrocentric chromosomes, No. 23 and 24. ...The litter size reduction amounts to 10 % , and it is supposed to be caused by the above mentioned meiotic disturbance.

Abb. 26: Weiße und blaue Rasse des Polarfuchses Alopex lagopus. Aus Eigener 1982.
Vernünftigerweise haben Christensen und Pedersen die beiden Chromosomenrassen nicht nicht in den Rang eigener Arten erhoben.
Würde jemand aufgrund der herabgesetzten Fertilität die beiden Chromosomenrassen dennoch in den Rang eigener Arten erheben, dann ginge hier die Artentrennung mitten durch die Blaufuchspopulationen. Das würde weiter bedeuten, dass bestimmte Blaufuchspopulationen mit dem Weißfuchs (in diesen beiden hauptsächlichen Rassen kommt der Polarfuchs vor) eine Art bilden würden, nicht aber mit den übrigen Blaufuchspopulationen. Man kann nur hoffen, dass niemand auf diese Weise zur Verwirrung in der Systematik beiträgt.
Auch bei Halbaffen (Prosimiae) und Affen (Anthropoidae) sind ausgeprägte Chromosomenpolymorphismen vorhanden. In einer Population der Gerboise-Affenart Tataryllus gracilis sind am gleichen Biotop Männchen mit 37 und Weibchen mit 36 Chromosomen gefunden worden, bei anderen dagegen mit 23 bzw. 22 Chromosomen. Nachkommen aus Kreuzungen der stark verschiedenen Chromosomenrassen sind steril (Grassé 1977, Hinweis von Schmidt 1985). Da die morphologischen Unterschiede fehlen, hat man auch von "kryptischen Arten" gesprochen. Man wende dieses Artkonzept nur einmal konsequent auf sämtliche Chromosomenrassen und -varianten aller Lebensformen auf unserer Erde an, um anhand der unübersichtlichen Konsequenzen zu verstehen, dass ein solcher Ansatz nicht weiter führt.
Von den nur in Afrika beheimateten Meerkatzen schreibt Mohr 1972/1982, p. 412:
Es gibt davon eine riesige Fülle sehr verschiedener Größe, die einander im Bau und Skelett sehr ähnlich sind, so daß man über ihre Artzugehörigkeit meistens nur nach Farbmischungen und Farbmustern zu ermitteln versuchen kann. Es ist also kein Wunder, daß die Systematiker noch keineswegs Einigkeit erzielt haben, wie und wo die Arten gegeneinander abzugrenzen sind, und daß De Vore feststellen konnte: "Wie viele Arten von Meerkatzen es tatsächlich gibt, hängt davon ab welcher Spezialist sie zählt."
Interessant ist in diesem Zusammenhang, dass nach den gründlichen Untersuchungen Chiarellis (1968) nicht nur abweichende Chromosomenzahlen zwischen den 'Arten' gefunden wurden (54 bis 72), sondern auch innerhalb der 'Arten', und das obwohl nur jeweils 2 bis 5 Tiere von den einzelnen Spezies untersucht wurden. Bei Cercopithecus l'hoesti wurden 58 und 60 Chromosomen festgestellt, bei C. neglectus 58 und 62, bei C. mona 66 und 68 und bei C. nicitans 66 und 70 (das Geschlecht wurde jeweils berücksichtigt; solche Unterschiede wurden z.B. auch zwischen zwei Männchen oder zwei Weibchen derselben Art festgestellt). Da diese Untersuchungen in der Regel an Individuen derselben Population einer Spezies vorgenommen wurden, darf wohl die Frage erhoben werden, ob mit den bisherigen Ergebnissen nicht erst die Spitze des Eisbergs zur Chromosomenvariabilität innerhalb der Arten gesehen wurde.
Es dürfte aber bereits klar sein, dass ein chromosomal definierter Artbegriff zur Trennung gleichartiger Populationen in verschiedenen Arten führen würde, - wozu man wohl nichts weiter sagen braucht.
Es gibt keine klare Korrelation zwischen der Chromosomenzahl und der morphologisch-anatomischen Divergenz. "So zeigt z. B. der Halbaffe Galago senegalensis nach H. LÜERS und Mitarb. (1974) (dort Originalliteratur) 38 Chromosomen, sein naher Verwandter G. crassicaudatus 62" (Schmidt 1985, p. 36). - Sowohl die Zahl als auch die Chromosomenmorphologie variieren innerhalb und zwischen den Arten so stark, dass jeder Versuch eines chromosomal motivierten Artbegriffs zu den unglaublichsten Konsequenzen führen würde. Die beiden eben zitierten Galago-Arten wären zum Beispiel um ein Vielfaches weiter voneinander entfernt als der Mensch (46 Chromosomen) von der Familie der Pongidae (Gorilla, Pan und Pongo mit ihren 48 Chromosomen), wie umgekehrt die Divergenz zwischen den oben aufgeführten Individuen innerhalb gleichartiqer Populationen von Meerkatzenarten zum Teil doppelt so groß wäre wie zwischen Menschen und Menschenaffen.
Wenden wir uns nun einigen Beispielen aus der Insektenwelt zu, um dann auf das Pflanzenreich zurückzukommen.
Barton spricht (1980, pp. 47-59) über "The Fitness of Hybrids between two Chromosomal Races of the Grasshopper Podisma pedestris" und kommentiert u.a.:
Genetic incompatibilities between two races of the grasshopper Podisma pedestris, which differ by a Robertsonian Fusion involving the sex chromosome, have been investigated ... F1 hybrids ... showed markedly reduced hatching success ... findings suggest that the karyotypic difference is only a weakly selected marker for more fundamental changes in genotype.
Ähnlich Barton und Hewitt (1981a und 1981b), wo die genetische Inkompatibilität weiter betont wird, weshalb man dieses Beispiel auch zum Thema des Gen-bedingten F2-Zusammenbruchs zitieren könnte. Die Autoren erheben die beiden Rassen jedenfalls nicht in den Rang eigener Arten, obwohl sie (1981a, p. 381) im Sinne des Neodarwinismus meinen, dass vielleicht viele weitere nicht entdeckte Hybridzonen scheinbar gleichförmige Gruppen in werdende Arten ("nascent species") aufteilen. Das letztere setzt nun wieder die ganze neodarwinistische Evolutionstheorie voraus, und wir haben ja oben schon zahlreiche Tatsachen und Einwände gegen diesen Ansatz zitiert, so dass wir an dieser Stelle nicht weiter auf diesen Punkt einzugehen brauchen.
Shaw und Wilkinson (1980), Moran, Wilkinson und Shaw (1980), Coates und Shaw (1982) und Shaw, Wilkinson und Coates (1982) gehen auf reproduktive Isolation und die Hybridzone zweier chromosomaler Rassen der Heuschrecke Caledia captiva ein. Shaw und Wilkinson schreiben 1980, p. 1:
These two taxa differ by a series of pericentric rearrangements which involve seven or eight members of the genome (2n = 11II + X0/XX). ... The F1 generation does not differ from the parental taxa in its fertility or viability. However, the F2 generation is completely inviable and in backcross generations viability ranges from 0 to 50 %.
(p. 24 zu den chromosomalen Veränderungen:) No obvious phenotypic effects result from these changes. Morphological divergence between the two taxa has therefore been minimal (Moran, 1978), life history parameters are indistinguishable and there are no overt differences between the habitats adjacent to the hybrid zone now occupied by these chromosomal taxa. Thus, we cannot define any selective agents which may be responsible for the observed chromosomal divergence.
In den 1982er Arbeiten (siehe oben) gehen die Autoren auf die chromosomale Komponente der reproduktiven Isolation ein. Ich möchte an dieser Stelle nur wieder festhalten, dass die Verfasser diese beiden, postzygotisch stark voneinander isolierten Rassen keineswegs in den Rang eigener Arten erhoben haben.
Suomalainen und Brown berichten (1934, pp. 170-176) über die Variation der Chromosomenzahl beim SchmetterLing Philaethria dido u.a.:
(p. 171:) We observed at least eight haploid chromosome numbers, namely 12, 12 large and 19-25 very small chromosomes, 21, 26 (needing confirmation), 51-52, 66-67, 72, and 87-88 (Fig.1).
Die Autoren weisen darauf hin, dass es sich hier nicht um Polyploidie handelt. Die DNA-Menge bleibt gleich. (Es handelt sich wahrscheinlich um Chromosomenumbauten, insbesondere Brüche und Fusionen.) Besonders interessant für unsere Frage nach dem Artbegriff ist die Tatsache, dass ganz verschiedene Chromosomenzahlen gleichzeitig bei Populationen an vier verschiedenen Orten gefunden wurden. Die Autoren schreiben (p. 173):
It would be interesting to know whether these sympatric individuals that have different chromosome numbers can hybridize. We do not find any evidence of individuals with meiotic features suggesting that there are hybrids between individuals with two different chromosome numbers. All individuals studied by us showed regular bivalent formation at meiosis. One might have assumed that this material would have contained at least one hybrid individual if chromosomally different individuals hybridized freely.
Zum Thema "systematische Schlussfolgerungen" stellen die Autoren fest:
Fig. 4 (hier als Abb. 27 p. 155 wiedergegeben) shows the subtle phenotypic differences, which may be seen in the wing pattern between Ph. dido with different chromosome numbers (12 + supernumb., 20, 52, 67, 88).
... There would seem to be three to five separable species in the "Ph. dido" complex (Brown 1981) in South America, and we expect at least one more in the Transandean region (México to western Ecuador). These will be formally named in a paper in preparation.
Sieht man sich die Abbildung dazu näher an, dann fällt einem auf, wie gering die Unterschiede zwischen den verschiedenen Linien sind. Die ventrale Oberfläche unterscheidet sich von der dorsalen ein- und derselben Linie oft stärker als diese Oberflächen zwischen verschiedenen Linien. Wenn man im übrigen bedenkt, was bei Schmetterlingen alles an Modifikationen möglich ist (vgl. p. 156) und dazu noch den stark ausgeprägten Geschlechtsdimorphismus bei manchen Arten berücksichtigt (p. 157), dann wird einem klar, dass die Tendenz, hier bis zu 6 verschiedene Arten zu unterscheiden, kaum mit den subtilen Unterschieden, "which may be seen in the wing pattern" (siehe oben) morphologisch motiviert sein kann. Bleiben die chromosomalen Unterschiede - und wegen der oben schon ausführlich beschriebenen Schwierigkeiten bei konsequenter Durchführung eines chromosomal begründeten Artbegriffs, ist zu 'befürchten', dass die Autoren hier tatsächlich mehrere Arten aufstellen werden. Mir erschiene es viel sinnvoller, wenn man in solchen Fällen - statt neue Arten aufzustellen - die chromosomalen Differenzen in Klammern hinter dem Artnamen mit geographischer Angabe vermerken würde. Die tertiäre Nomenklatur wäre wohl nur im Zusammenhang mit stärkeren morphologischen und ökologischen Unterschieden angebracht.

Abb.27: Representative voucher specimens of Philaethria for which chromosome numbers have been determined. v = ventral wing surface, d = dorsal, with left side given first.
A Philaethria pygmalion, Humaitá, Amazonas, Brazil. n = 29 (v/d). B Ph. wernickei, Campinas, Sao Paulo, Brazil, n = 29 (d/v). C-M All belong to Ph. dido complex: C Quinze de Novembro, Serra do Cachimbo, Pará, n = 88 (v/d). D Tefé, Amazonas, Brazil, n = 88 (v). E Belém, Pará, n = 88 (v). F Linhares, Espírito Santo, n = 51 (or 67) (v/d). G Igarapé Santa Julia, km 1,135 of Cuiabá-Santarém road, Pará, n = 52 (v). H Linhares, Espírito Santo (female), n = about 20 (v). I Above Santa Ana, Táchira, Venezuela n = 52 (d/v). J La Fundación, Táchira, Venezuela, n = 72 (or 21), (v/d). K 27 km S of El Manteco, Bolivar, Venezuela, n = 12 + 19-25 supernumeraries (v/d). L Salinas, Pará, n = 12 + 19-25 supernumeraries (or 52) (v). M Soure, Ilha de Marajó, Pará, n = 12 (v). Colours are black und green for dorsal surface, and black, green, silver and reddish für ventral. Additional phenotypes, possibly with further numbers, have been examined from north of Manaus, Amazonas in Brazil, and from Barinitas, Venezuela; Cali, Colombia; Panamá Canal Zone; and Costa Rica. Aus Suomalainen und Brown 1984.
Über die Biotope der derzeit mit Artnamen versehenen verschiedenen Linien Philaethria dido, Ph. pygmalion und Ph. wernickei schreiben die Autoren (p. 175):
Philaethria dido is a sun-loving species, which inhabits the canopy, clearings, and edges of rain forests over an area that extends over most of the neotropics (southern México to southeastern Brasil). Philaethria pygmalion is restricted to non-forest habitats from eastern Columbia (Villavicencio) and southern Venezuela (Yavita) south to central Brazil and down the Amazon to near its mouth; the closely related Ph. wernickei occurs along the eastern coast of Brazil from Rio Grande do Norte south into Uruguay and northern Argentina.
Die morphologischen Unterschiede sind allerdings so schwach (vgl. Abb. 27), dass man die Frage stellen könnte, ob denn die hier vorliegenden ökologischen Unterschiede ausreichen, um überhaupt nur einmal die tertiäre Nomenklatur für diese "Arten" einzuführen. Bei der oben zitierten Tabakmaus gibt es wenigstens ausreichende morphologische Besonderheiten, die in Verbindung mit chromosomalen Abweichungen die tertiäre Nomenklatur rechtfertigen. Was machen denn unsere Systematiker, wenn Philaethria pygmalion (n = 29) mit Ph. wernickei (n ebenfalls = 29) kreuzbar ist und diese beiden Arten vielleicht noch mit chromosomal passenden dido-Linien kreuzbar sind? Die chromosomal motivierte Arttrennungslinie würde den Artbegriff an anderen Stellen wieder aufheben und eine konsequente Anwendung eines solchen Artbegriffs würde (und hat schon) zu zahlreichen Schwierigkeiten in der oben beschriebenen Weise geführt.
Weder chromosomal noch durch die geringen Unterschiede im Muster der Flügel ist eine Auftrennung in verschiedene Arten überzeugend. Der folgende kleine Abstecher zur Frage, was an Muster- und Farbunterschieden bei Schmetterlingen innerhalb der Art möglich ist, soll den letzteren Punkt noch weiter verdeutlichen.
So berichtet Wigglesworth 1971, p. 109:
Wenn bei einem Insekt mehr als eine Generation pro Jahr auftritt, gibt es manchmal Unterschiede zwischen der Frühlings- und der Sommergeneration in Struktur und Aussehen. Eines der berühmtesten Beispiele ist das Landkärtchen Araschnia 1evana (Nymphalidae), dessen blasse Frühlingsform levana ursprünglich als eigene Art angesehen wurde, ganz verschieden von der Form prorsa, einem dunkel gefärbten Insekt, das im Spätsommer erscheint. Sie sind in Wirklichkeit Saisonformen einer einzigen Art: Die Form levana entsteht aus einer Diapausenpuppe, die aus den kurzen Tagen des vorhergehenden Herbstes stammt, die Form prorsa aus einer Puppe ohne Diapause. Indem man die Raupen einem langen oder einem kurzen Tag aussetzt, kann willkürlich jede der beiden Formen zu jeder Jahreszeit erzeugt werden.
Stanek beschreibt 1977, p. 86 diesen Fall anhand mehrerer farbiger Abbildungen und führt weitere Beispiele auf:
Der Netzfalter Araschnia levana L.
ist der kleinste und einer der schönsten echten paläarktischen Fleckenfalter. Diese eurosibirische Art lebt in Laubwäldern und ist ein weiteres europäisches Beispiel für die jahreszeitlich abhängigen Veränderungen bei Schmetterlingen. Zwei Generationen, die in verschiedenen Jahreszeiten geboren werden, sind so unterschiedlich, daß sie für zwei besondere Arten gehalten werden können, was auch geschah, indem sie unterschiedlich beschrieben und benannt wurden. In den Tropen tritt dieser Fall häufig ein. So ist zum Beispiet Precis octavia Crammer während der Trockenzeit im ganzen dunkelblau und während der Regenzeit rot. Die Zwischengeneration ist halb blau und halb rot gefärbt. Diese Verändenmgen bereiten den Entomologen manchmal Schwierigkeiten.
Details zu weiteren Beispielen und zahlreiche Abbildungen, vgl. die oben zitierten Autoren sowie Dierl (1981), Forster und Wohlfart (1960-1981) und Novak und Severa (1983).
Ein Beispiel für Geschlechtsdimorphismus sei mit dem folgenden Beispiel vor Augen geführt. Die Unterschiede innerhalb der Art können erstaunlich sein (aus Stanek p. 32) (Philaethria-Unterschiede verblassen dagegen fast ganz):
Papilio glaucus L.
ist eine der großen Arten der Ritterfalter. Ihre Heimat sind die gemäßigteren Gebiete Kanadas und die mittleren und östlichen Länder der Vereinigten Staaten bis nach Texas und Florida. Hier leben mehrere Formen. Bei den interessantesten sind beide Geschlechter unterschiedlich gefärbt. Die Männchen der südlicheren Formen (rechts) tragen die bei den Ritterfaltern üblichen Farben - kanariengelb und samtschwarz mit orangefarbenen und blauen Abzeichen auf den Hinterflügeln. Die Weibchen (links) sind dagegen schwarzbraun und haben reichen blauen und rötlichen Schmuck auf den Hinterflügeln.

Abb. 28: Aus Stanek 1977. S.V. +- 95 mm. N. R. Liriodendron, Betula, Prunus, Fraxinus,Tilia, Salix, Populus, Sorbus, Malus, Acer, Magnolia.
Kommen wir auf die Frage nach der Bedeutung chromosomaler Unterschiede zurück.
Zahlreiche weitere Beispiele für Chromosomenverschiedenheiten innerhalb von Arten sind bekannt. Foresti et al. (1984) berichten über den Fisch Gymnotus carapo, dass die Chromosomenzahl von Individuen von fünf verschiedenen Lokalitäten zwischen 2n = 42 bis 2n = 54 varierte.
Mit solchen Befunden steht die leichte Induzierbarkeit von Translokationen bei vielen Spezies in Einklang. So berichten z.B. Robinson und van Heemert (1981), dass sie nach Einsatz von Röntgen- und Neutronenstrahlen bei der Zwiebelfliege 61 Translokationen isolieren konnten (Fertilität zwischen 40-70 %).
Suguna et al. erwähnen für ihre strahleninduzierten perizentrischen Inversionen und Translokationen bei Anopheles albimanus folgende Anwendungsmöglichkeit (1981, p. 57):
Assays of the partial sterility inherent in male heterozygotes for a pericentric inversion and a male-linked translocation indicated that the combination of inversions and translocations could be used in genetic control schemes.
Nach morphologischer Klassifizierung ist das Genus Allium allein in Griechenland mit mehr als 50 Spezies vertreten. Tzanoudakis berichtet zu seinen Karyotyp-Studien an 10 Taxa von Allium, Section Scorodon aus diesem Land u.a. (1983, p. 281):
The results of the present study have shown that similarities between the karyotypes of several species may exist and sometimes they confirm the relationships of the species as suggested by morphological characters, but the opposite is not always the case. Karyotype differences, of structural or numerical nature, may not have any taxonomical value especially when they are observed in species indicating remarkable chromosome morphological variation.
Die Beschreibung aller chromosomal unterschiedlichen Linien als eigene Spezies ist in der Botanik wahrscheinlich mit der Schwierigkeit verbunden, dass solche Linien schneller entstehen, als die Systematiker sie klassifizieren könnten.
Über die komplexe Zusammensetzung der Spezies Scilla scilloides in Korea berichtet Araki (1985, p. 3):
This species has been shown to be composed of different cytogenetic types, i.e. AA (2n = 16), BB (2n = 18), A BB (2n = 26), BBB (2n = 27), AA BB (2n = 34), A BBB (2n = 35), BBBB (2n = 36) and AA BBB (2n = 43), where A and B denote genomes with x = 8 and x = 9 respectively (...).
(pp. 5/6:) Usually, AA was found isolated from the other types growing in relatively undisturbed sites in hilly regions. In contrast, AA BB tended to grow in ruderal and disturbed habitats. However, coexistence of AA with AA BB within a population was found at Nam Weon (Jeju Island) and in Seoul. These cases probably might be due to recent disturbances of habitat by man. In these mixed populations, there was no evidence of natural hybridization between AA and AA BB. This is evidently due to flowering time of the tetraploid AA BB being later than that of diploid AA...
(p. 8:) The diploids AA and BB are allopatric in distribution. With different flowering times, they can be hybridized only by artificial means (...).
Araki hat aus diesen cytologischen und ökologischen Differenzen vernünftigerweise keine Serie neuer Arten kreiert.
Nazeer und Khoshoo listen für Sansevieria cylindrica folgende Chromosomenzahlen auf (1984, p. 326): 2 n = 18 (Sarkar et al. 1977; 40 (Roy, M. 1956); 84 (Roy, R.P. und Misra 1961); 92 (Sharma und Chaudhuri 1964); 102-104 (Heitz 1926); 112 (Nazeer und Khoshoo 1984); 120 +- (Menzel und Pate 1960).
Sau und Sharma stellen (1983, p. 370) fest, dass intraspezifische Chromosomenrassen bei den Orchidaceae häufig vorkommen.
Gottschalk schreibt 1976, p. 95:
Die Vorstellung, eine pflanzliche Species habe eine ganz bestimmte Chromosomenzahl, muß bei Berücksichtigung der in den letzten Jahrzehnten erarbeiteten Befunde als unbegründet betrachtet werden. Schon im Jahre 1933 hat DARLINGTON eine Liste mit 33 Arten publiziert, für die jeweils mehrere Chromosomenzahlen bekannt waren. Wenige Jahre später führt TISCHLER (1935) etwa 80, MÜNTZING (1936a) 70 Arten mit intraspezifischer Polyploidie an. Auch FAGERLIND (1937) hält das Phänomen auf Grund seiner Untersuchungen an Rubiaceen für weit verbreitet. Der Chromosomenatlas von DARLINGTON und WYLIE (1961) gibt für zahlreiche Arten mehrere Chromosomenzahlen an, die sich häufig, aber durchaus nicht immer in eine polyploide Reihe aus euploiden Valenzen einordnen lassen. Oft sind darüber hinaus noch mehrere aneuploide Zahlen bekannt geworden. Die im Pflanzenreich auftretende Vielfalt der Chromosomenzahl und Genomstufen ist erst zu einem kleinen Teil erfasst. Ihre weitere Bearbeitung wird uns interessante Einblicke in die Vielfalt phylogenetischer Entwicklungsprozesse geben, die wir zur Zeit nur in groben Umrissen überblicken können. In den Tab. 5 und 6 des vorliegenden Buches sind knapp 200 Arten aufgeführt, für die neben der diploiden Valenz noch eine oder mehrere polyp
loide Genomstufen bekannt sind, während die Tab. 7 entsprechende Arten mit aneuploiden Serien enthält.Die Tabellen umfassen allein sieben Seiten und erheben dabei noch keineswegs den Anspruch auf Vollständigkeit (der an Details interessierte Leser sei auf Gottschalks Arbeit verwiesen).
Derselbe Verfasser weist (1984, p. 270) darauf hin, dass "mehr als 1000 Arten bekannt (sind), in denen neben der diploiden Ausgangsform polyploide Rassen unterschiedlicher Valenzen feste Bestandteile bestimmter Pflanzengesellschaften sind" und fährt fort (pp. 270/271):
Es handelt sich hier nicht nur um polyploide Reihen mit den Valenzen 2n, 3n, 4n, 6n, 8n oder ähnliche Kombinationen. Nicht selten sind daneben auch aneuploide Chromosomenzahlen vertreten. Auf diese Weise kann bei der gleichen Art eine außerordentlich hohe Anzahl verschiedener Chromosomenzahlen zustande kommen. So sind von der Liliacee Allium nutans bisher mehr als 60 Chromosomenzahlen zwischen 2n = 16 und 108 bekannt.
Die intraspezifische Polyploidie ist im Planzenreich so verbreitet, daß man nicht mehr von der früheren Vorstellung ausgehen kann, eine jede Art sei durch eine einzige Chromosomenzahl gekennzeichnet.
Angesichts dieser Tatsachen dürfte klar sein, dass der Versuch, den Artbegriff chromosomal zu definieren und spätestens bei herabgesetzter Fertilität zwischen zwei Linien von zwei verschiedenen Arten zu sprechen zum Scheitern verurteilt ist (abgesehen davon, dass die Fertilitätsfrage bei vielen sich vegetativ fortpflanzenden Arten - was zum Beispiel auf viele Allium-Linien zutrifft - sowieso schwer zu beantworten ist).
Andernfalls müssten Tausende (oder sogar Hunderttausende!) neuer chromosomaler Arten aufgestellt werden. Das wiederum würde in ähnlicher Weise, wie für den morphologischen Artbegriff oben ausgeführt, zu einer uferlosen Speziesmacherei führen und schließlich soviel Unübersichtlichkeit und Unordnung schaffen, dass der Zweck jeder systematischen Arbeit verfehlt wäre. (In Klammern sei noch erwähnt, dass sich viele der chromosomalen Rassen morphologisch nicht voneinander unterscheiden, was wohl Systematiker bisher mit dazu bewogen hat, solchen Rassen keinen Artstatus zuzuschreiben.)
Oben haben wir mit Beispielen wie den Säugetiergattungen Mus, Sorex, Alopex, den Insektengattungen Podisma, Caledia, Philaetria und Beispielen aus dem Pflanzenreich auf chromosomale Unterschiede innerhalb der Arten hingewiesen (vgl. auch Herre und Rohrs, zitiert Seite 33). Es sei weiter hervorgehoben, dass solche chromosomalen Unterschiede keineswegs gleichmäßig auf die verschiedenen Tier- und Pflanzengruppen verteilt sind. So meint z.B. Imai (1983, p. 1160) nach Untersuchungen an 237 Genera der Säugetiere, dass hier "speciation without karyotype alternation" dominiert (wobei der neodarwinistische Artbegriff impliziert wird.)
Die Frage nach dem Grund der ungleichmäßigen Verteilung findet ihre Antwort z.T. in den Chromosomenstudien von Lima-de-Faria. Der Verfasser beschreibt 1979, pp. 27-28 die herkömmliche Auffassung von der Chromosomenstruktur im Gegensatz zu seiner eigenen (auszugsweise) wie folgt:
The current view of the chromosome is that it is a chaotic structure in which genes, mutations and structural rearrangements occur at random (e.g. White 1973). In opposition to this view I have proposed the concept of the chromosome field (Lima-de-Faria 1954, 1976a).
The chromosome appears to be a hierarchic structure in which centromeres and telomeres are the main organizers. Each gene has an optimal position within the centromere-telomere field. This position is called the gene territory. It is postulated that the gene has sensors that recognize messages from the centromere and the telomere regions.
Über die Verteilung der Gene für die 28 S und 18 S ribosomale RNA (Nukleolus Organisator Region) stellt er fest (p. 29):
The distribution of these genes is so regular that it can be predicted by means of a straight-line equation (Lima-de-Faria 1973).
Auf Seite 35 weist er noch darauf hin, dass die konservativen Chromosomenumbauten, d.h. diejenigen, die das Chromosomenfeld aufrecht erhalten, auch diejenigen sind, die am häufigsten in natürlichen Populationen vorkommen.
In seiner großen Arbeit von 1983 (MOLECULAR EVOLUTION AND ORGANIZATION OF THE CHROMOSOME; 1186 Seiten) hat der Verfasser das Thema eingehend behandelt. Zur neodarwinistischen Position meint er u.a. (p. XIII): "One resorts to natural selection every time one lacks knowledge of the molecular mechanisms involved." (p. 329:) "According to this view most mutations occur at random and little or nothing is predictable concerning the genetic behaviour or the evolution of the chromosome. Chromosome evolution is assumed to be mainly dictated by natural selection (White 1973, Ohno 1974b), which is used as an 'explanation' to cover up for the lack of molecular evidence." (p. 1023:) "What I am trying to convey is that due to the absence of knowledge of molecular mechanisms, selection has been employed like a kind of general remedy by the biologist. Every time a phenomenon appeared in biology, and one obviously ignored its mechanism, selection was invoked as an explanation and the matter was settled."
Im Kapitel 16 (pp. 329-417), das mit dem Thema "Disorder in biological phenomena is synonymous with ignorance" eingeleitet wird, kommt Lima-de-Faria ausführlich auf das Chromosomenfeld zu sprechen. Ohne auf die große Zahl von Untersuchungen und Argumenten einzugehen, die für eine hierarchische Organisation der Chromosomen sprechen, seien seine Definitionen des Chromosomenfeldes und die damit für unsere Fragestellung relevanten genetischen Konsequenzen zitiert. Da seine Auffassung noch nicht die ihr gebührende Beachtung zuteil geworden ist, zitieren wir den Autor etwas ausführlicher.
Der Verfasser schreibt 1983, pp. 377-378:
The order within the eukaryotic chromosome is much more evident and widespread than could have been expected a few years ago. The definition of the field which was originally given was as follows (Lima-de-Faria 1954).
"The expression chromosome field (vom Verfasser kursiv, wie auch in den folgenden Ausführungen) refers to the fact that the variation observed within the gradients takes place in such a way that the properties exhibited by certain entities in one part of the chromosome bear a definite relation to the properties exhibited by other entities in other parts of the chromosome. These relations are such that the different entities appear to share in the formation of the chromosome pattern with unequal importance. Whereas kinetochores appear to be primarily responsible for the formation of the gradients, chromosome ends, knob formations and their adjacent regions, recurrent seriations and chromomeres or fibrillae apparently interfere with the formation of the gradients on a successively smaller scale. Moreover - like the embryological field - the chromosome field can maintain its pattern when its mass is either reduced or increased, that is to say, when the arm length decreases or increases."
At that time the definition was based solely on the information from the chromomere gradients. One can see that all the results which were added since then have not changed it in any way but have only reinforced the evidence.
More recently I redefined it (Lima-de-Faria 1980a). 'The chromosome field is defined as the system of interrelationships, existing between the different DNA sequences, which results in the establishment of the order and hierarchy observed within the centromere-telomere segment'.
The basic principle which emerges from this concept is that it is the relative position of each DNA sequence within the unit built by the centromere-telomere segment which is decisive in determining the permanence and the function of the sequence.
Mit dem Konzept des Chromosomenfelds sind zahlreiche genetische Konsequenzen verbunden, die der Verfasser auch im einzelnen in seiner Arbeit diskutiert. Wir wollen an dieser Stelle wieder nur die für den Artbegriff wichtigen Punkte der chromosomalen Umbauten zitieren. Lima-de-Faria nimmt folgende Einteilung der chromosomalen Umbauten vor (p. 378):
Rearrangements are classified as conservative, discordant, disruptive, destructive and incompatible due to whether the genes maintain the same order within the field or this order is successively changed to result in the destruction of the field.
Auf den Seiten 388 - 403 geht der Autor auf die daraus folgende Klassifikation der Translokationen, Inversionen, Duplikationen und Deletionen ein. Da wir in unseren Beispielen desöfteren schon den Translokationen begegnet sind, seien diese im folgenden weiter zitiert (pp. 388, 392-394):
Classification of translocations. Conservative translocations are of three types. Reciprocal symmetrical centric fusion (Robertsonian), and dissociation. The changes in gene order that they produce in relation to the centromere and to the telomere are minimal (Table 16.12 [Die Tabelle umfasst allein bereits zwei Seiten. Für das Verständnis sind vor allen die Abbildungen wichtig, die wir auf den folgenden Seiten wiedergegeben haben.] and Fig. 29.1.).
Discordant translocations have a moderate effect on the field. There are two types: intrachromosomal involving a small shift; and interchromosomal, involving an eucentric small transposition (Fig. 29.2.).
The disruptive translocations are of five types and have a drastic effect on the field. Intrachromosomal, large shift; interchromosomal, eucentric large transposition; interchromosomal, dyscentric transposition; tandem fusion involving two acrocentrics; and tandem fusion involving one acrocentric and one metacentric. The first type moves telons to a centron territory and vice versa (Fig. 29.3.) In the second type centrons become telons and other genes change position. Telons become centrons in the third type. Medons become telons and vice versa (fourth type). The same situation occurs in the fifth type (Fig. 29.4.). The position of the genes within the field is drastically changed.
The destructive translocations create lethal effects for their chromosomes. A reciprocal asymmetrical with distal breaks produces a dicentric and a fragment without centromere. A reciprocal asymmetrical with proximal breaks has the same effect (Fig. 29.5.). When the centromeres become located close to each other, one is usually suppressed restoring the chromosome field (Niebuhr 1972).
The incompatible type is the one in which a single break on one chromosome not followed by a break in another leads to the formation of (1) one chromosome without telomere, which due to the lack of this necessary segment for field action cannot survive, (2) a free telomere segment without centromere in which the field is equally disrupted, and (3) a complete chromosome, which due to the fact that its end is saturated by a telomere cannot attach itself to the free telomeric segment. It is thus not surprising, in the fact of the evidence from the chromosome field, that there is little or no experimental evidence for ihe occurrence of this type of translocation.
Under exceptional conditions the union of the normal telomere (r Fig. 29.5.) with a telomere in reverse position (h Fig. 29.5.) would create a double telomere with a symmetric configuration. This would not be terminal but nearly terminal. The presence of symmetrical telomeres has not been reported as far as I know and they are expected to occur only under exceptional conditions.
The terms used in the description of the different rearrangements such as eucentric, dyscentric, symmetrical, asymmetrical are part of the conventional terminology (cf. Rieger et al. 1968).
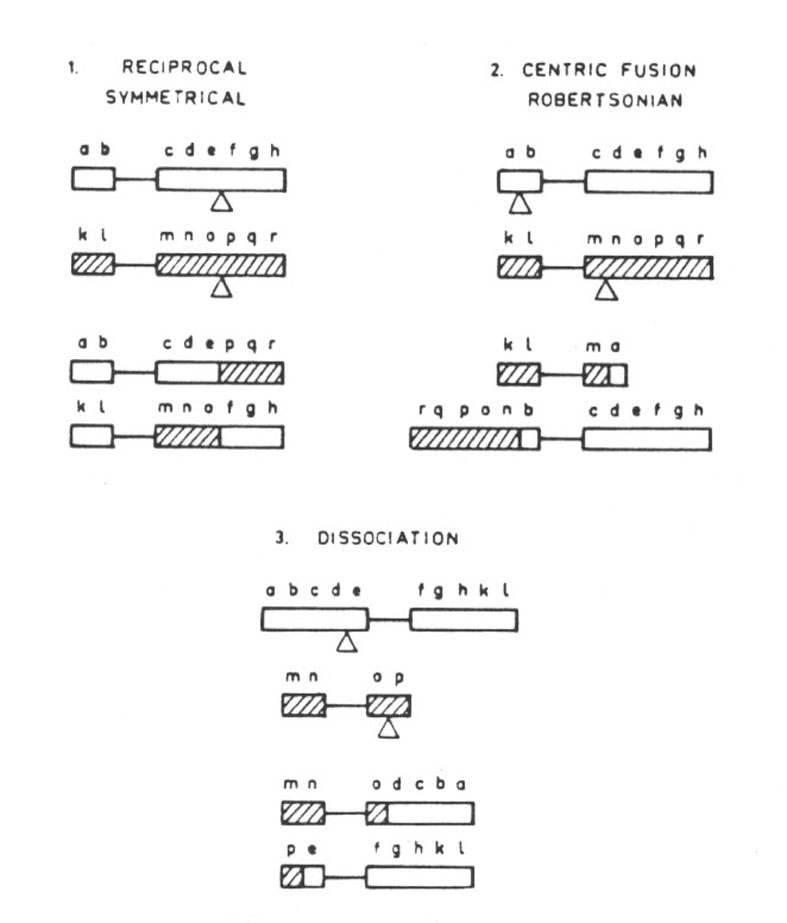
Abb. 29.1.: Classification of translocations according to the chromosome field. They may maintain the order previously established or they may disturb or destroy it. Translocations can, accordingly, be classified as Conservative, Discordant, Disruptive, Destructive or Incompatible. Schematic examples of these different situations are given. Conservative translocations: reciprocal symmetrical (1); centric fusion, Robertsonian (2); dissociation (3). Aus Lima-de-Faria 1980 und 1983/1986.
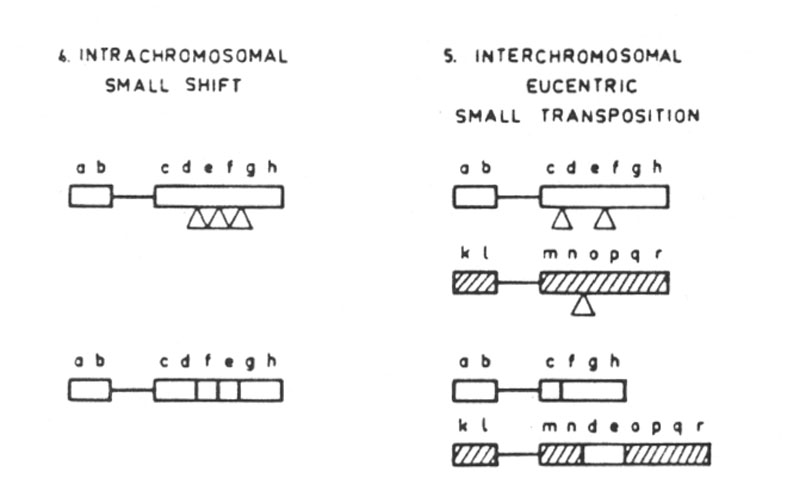
Abb. 29.2.: Discordant translocations: intrachromosomal, small shift (4); interchromosomal eucentric, small transposition (5). Aus Lima-de-Faria 1980 und 1983/1986.
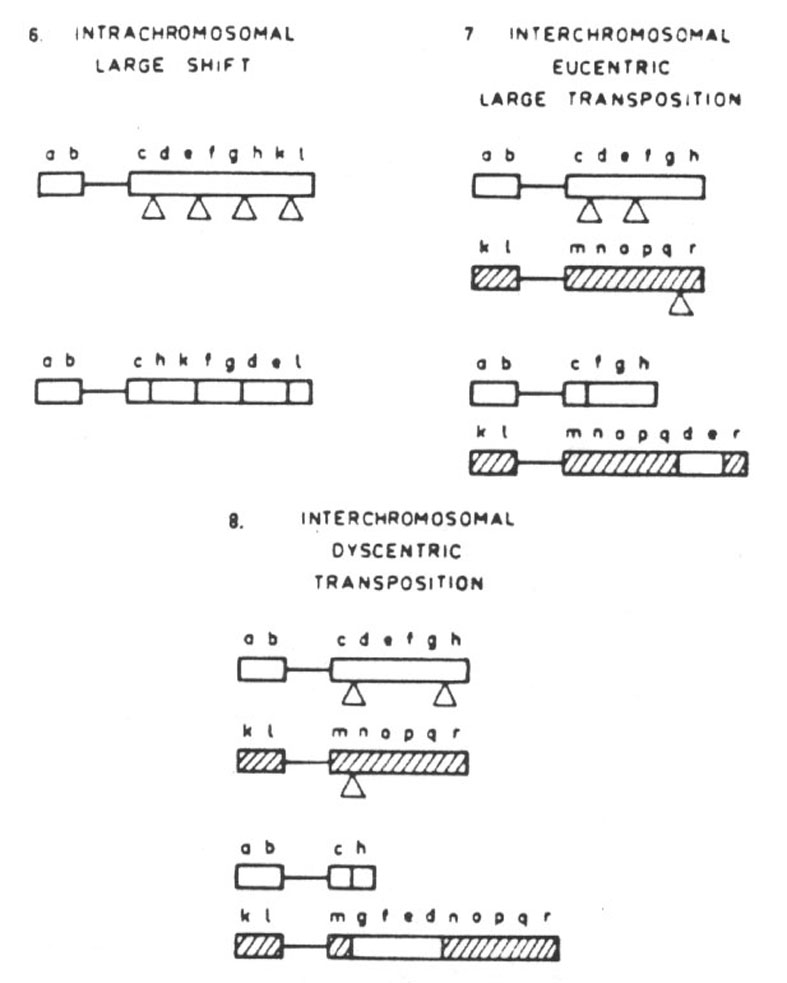
Abb. 29.3.: Disruptive translocations: intrachromosomal, large shift (6); interchromosomal eucentric, large transposition (7), interchromosomal dyscentric transposition (8). Aus Lima-de-Faria 1980 und 1983/1986.
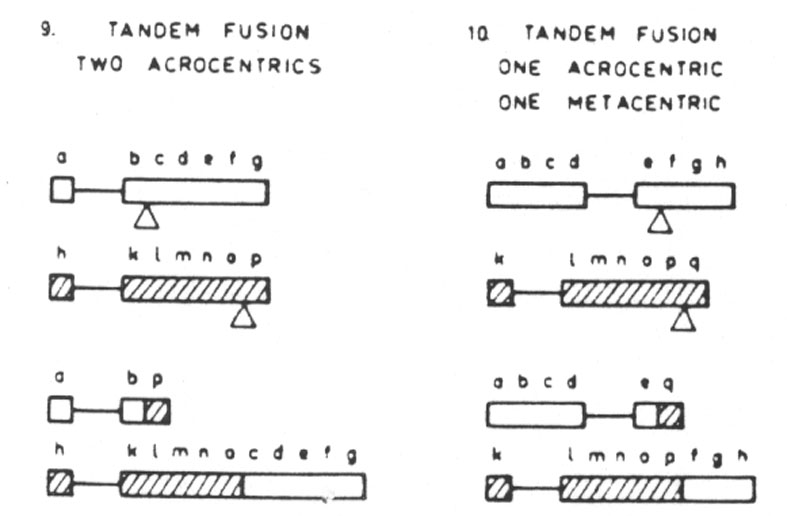
Abb. 29.4.: Disruptive translocations: tandem fusion, two acrocentrics (9); tandem fusion, one acrocentric and one metacentric chromosome (10). Aus Lima-de-Faria 1980 und 1983/1986.
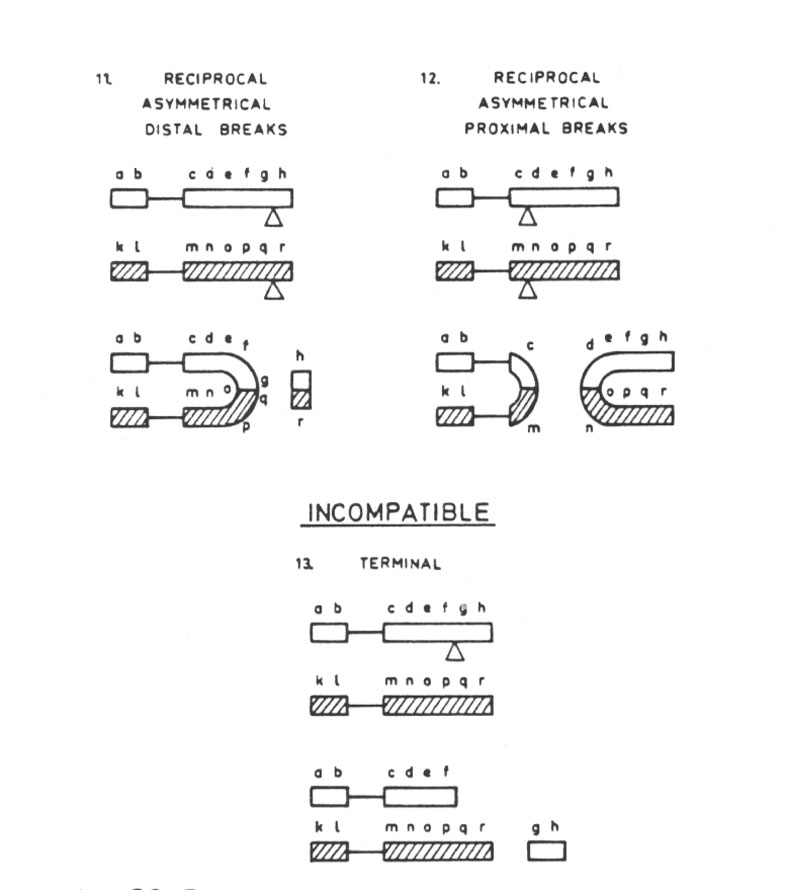
Abb. 29.5.: Destructive translocations: reciprocal asymmetrical with distal breaks (11); reciprocal asymmetrical with proximal breaks (12). Incompatible translocations: terminal (13). Aus Lima-de-Faria 1980 und 1983/1986.
Damit dürfte gezeigt sein, dass zahlreiche chromosomale Umbauten, die zu entsprechend unterschiedlichen, aber lebensfähigen Individuen und Populationen führen, nicht einfach zufällig, sondern 'chromosomal' determiniert sind. Die Möglichkeiten sind zwar sehr groß, aber dennoch begrenzt, womit Lima-de-Faria auch begründen kann, warum es (wie zitiert) die 'konservativen' Chromosomenumbauten sind, die am häufigsten in natürlichen Populationen vorkommen.
Wenn sich auch manche Details der Chromosomenfeld-Forschung in Zukunft noch anders darstellen dürften als heute von Lima-de-Faria konzipiert, so steht dieser Autor mit seinen Untersuchungen doch keineswegs allein da. Zahlreiche neuere Arbeiten zur Nicht-Zufälligkeit chromosomaler Brüche und Umbauten liegen vor.
Baverstock, Gelder und Jahnke haben (1983, pp. 93-103) bei australischen Ratten-Populationen festgestellt, dass die 'Chromosomenevolution' erstaunlich schnell vonstatten gegangen ist und sich vollkommen auf Robertsonsche Translokationen beschränkt.
Peters knüpft in seiner Arbeit THE RECURRENCE OF CHROMOSOME FUSION IN INTER-POPULATION HYBRIDS OF THE GRASSHOPPER Atractomorpha similis (1982) an die Ergebnisse Whites mit seinem Begriff der 'karyotypischen Orthoselektion' an, kommt jedoch zu einer anderen Ursache des Phänomens. Er erwähnt einleitend die Befunde von White und anderen und fasst dann nach Aufführung der eigenen Untersuchungen sein Anliegen zusammen. Beide Punkte seien im folgenden zitiert (pp. 323/324 und 345/346):
As pointed out by White (1975) there is a "tendency for similar structural changes to establish themselves in one member of the karyotype after another". He adds that "these conditions would not be found with such regularity if the structural changes which have become established in phylogeny were of all possible types". Spectacular examples of such "karyotypic orthoselection" (White, 1975) have been observed in a number of organisms and some evidence of it is apparent in a wide range of taxa (Swanson et al. 1981).
.....
As I have attempted to show in this paper however, karyotypic orthoselection relies fundamentally on the non-randomness of structural mutation. Chromosome breakage occurs at specific sites throughout the genome, to produce a predominance of one type of rearrangement. Natural selection therefore has little from which to choose. It may eliminate many rearrangements, but most of those that survive, as well as those which do not, will be of one type. Thus natural selection is not critical to karyotypic orthoselection as defined here.
In diesem Zusammenhang sind die Koppelungs-Homologien von großem Interesse. Da die Funktionsfähigkeit des X-Chromosoms für die männlichen Säugetiere in besonderem Maße gewährleistet sein muss, war hier von Ohno (1970) eine weitgehende Homologie für die ganze Klasse der Mammalia postuliert worden. McKusick schreibt zu diesem Punkt 1980, p. 379:
... absolute homology for gene content appears to exist for the X-chromosomes of all placental and marsupial mammals. Implied is that no evolutionary change has occurred in the mammalian X in nearly 100 million years.
- und schreibt dann zu den Autosomen des Menschen im Vergleich zu denen der Maus:
Less expectedly, extensive syntenic homology is being found in that more distantly related mammal, the mouse.
AVP (antiviral protein or interferon receptor) and SOD-1 (cytosolic superoxide dismutase), which are both on chromosome 21, the "Down syndrome" chromosome, in man, are also syntenic in the mouse, on chromosome 16.
Gc (group-specific component, the vitamin-D binding alpha-2-globulin) and serum albumin are closely linked in both horse and man, another example of evolutionary conservatism of linkage. The tight linkage of thymidine kinase (TK) and galactokinase (GalK) (on human chromosome 17) has been conserved in chimpanzee, chinese hamster, mouse and other species.
Darüber hinaus ist die Gc-Serumalbumin-Koppelungsgruppe auch beim Huhn gefunden worden (Juneja et al. 1982).
In ähnlicher Weise berichten Andersson et al. 1983, p. 361:
Linkage homologies also may be found in distantly related species as shown by the close linkage of the loci for albumin and serum vitamin D binding protein in chickens, horses and humans.
(p. 363 über ein größeres, konserviertes Chromosomensegment:) It is interesting from an evolutionary point of view that such a large chromosome segment appears to be conserved in three different mammalian species [eigntl. "families"], namely Equidae, Muridae, and Leporidae. The observation becomes even more interesting in view of the report of an extraordinary rapid chromosomal evolution in the family Equidae in comparison with other vertebrate families.
It is possible that a considerable portion of the homologous chromosome segment observed on equine LG II, mouse chromosome 8, rabbit LG VI, and rat LG V (Figure 2) also is conserved on chromosome 16 in humans. This concept is based, first, on the fact that the loci for adenine phosphoribosyltransferase (APRT) and mitochondrial GOT are syntenic on chromosome 8 in the mouse and 16 in humans. Furthermore, the possible existence of a more extensive homology between these chromosomes is indicated by the assignment to human chromosome 16 of the loci for carboxylesterase (ESB 3), a lipase (LIPB; a lipase is a type of esterase) and a chymotrypsin protease (CTRB). A chymotrypsin protease Locus, Prt 2, is located at an intermediate map distance between e and Es-1 on mouse chromosome 8. [Vgl. auch M. Münke und U. Francke (1987): The physical map of Mus musculus chromosome 11 reveals evolutionary relationships with different syntenic groups of genes in Homo sapiens. J. Mol. Evol. 25, 134 - 140.
Auffallende Übereinstimmungen in der Chromosomenstruktur gibt es auch bei so verschiedenen Vogelordnungen wie den Hühnern, Tauben und Möwen.
Carlenius et al. erwähnen u.a. folgende Punkte (1981, p. 66):
Stock et al. (1974) found a surprising correspondence between the pattern of the G-bands on the three largest chromosomes in three species of the two orders Galliformes and Columbiformes. In this work we can establish a G-band homology for the four largest chromosomes between Gallus domesticus and four Larus species (Charadriiformes) except for a short segment on the terminal part of the long arm on chromosome 1 which seems to be lacking in the Domestic Fowl, and the more pronounced centromeres on the Larus chromosomes (Ryttmann et al. 1979). As already pointed out by Stock et al. (1974), these findings present a problem in interpreting the relationship of karyotyped divergence to phylogeny in birds.
Presenting a contrast to this homology Stock et al. (1974) discuss the surprising difference between some close relatives of the chicken in morphology of chromosome 2. This lack of homology is confirmed by a study of G-banded chromosomes from three species of the order Galliformes (Ryttmann et al., in prep.).
Gäbe es kein Chromosomenfeld, dann dürfte es nach all den in die Milliarden gehenden, zufälligen Umstrukturierungen in den postulierten Jahrmillionen keine ausgedehnten Homologien in den Autosomen mehr geben.
Für diese Fragestellung sind weiter die Arbeiten von Gill et al. (1980), Thierry-Mieg (1982), Schmid et a1. (1982), Antonucci et al. (1984), Ohno (1979), Hotta et al. (1985), Womack und Moll (1986) u.v.a. von Interesse, da sie mit zahlreichen Details den Ansatz weiter fundieren.
Ergebnisse für den geordneten Ablauf der Ereignisse liegen aus den verschiedensten Bereichen der Chromosomenforschung vor. So berichten z.B. Raina und Rees in ihrer Arbeit über DNA VARIATION BETWEEN AND WITHIN CHROMOSOME COMPLEMENTS OF Vicia SPECIES (1983, pp. 335-346) folgendes (p. 343):
... we conclude from the evidence presented that the accretion (or diminuation) of chromosomal DNA by the amplification (or deletion) of base sequences within each complement affected each chromosome to the same degree. This despite the fact that there is a considerable variation in size between chromosomes within the Vicia complements. A similar pattern of DNA change is associated with the divergence and evolution of species in Lolium and Festuca (Seal and Rees, 1983) and in Lathyrus (Narayanan, 1982). The indications are that the pattern may be widespread arnong genera of flowering plants. The phenomenon is surprising. One would expect the mutational events leading to the amplification or deletion of DNA base sequences to affect the DNA molecules at random. One would expect, in consequence, that the accretion or diminuation of DNA among chromosomes within a complement would be proportional to chromosome size and DNA content.
Das alles ist jedoch nicht der Fall. Dennoch bleiben die Autoren bei einer neodarwinistischen Deutung durch zufällige Mikromutationen und Selektion, obwohl das Phänomen bei weiteren Pflanzengattungen festgestellt wurde und im Pflanzenreich weitverbreitet sein könnte. Ähnliche Genomunterschiede beim Flachs sind jedoch nicht auf diese Faktoren zurückzuführen, sondern wahrscheinlich auf Transposonaktivierung in Korrelation mit der gegebenen Chromosomenstruktur (vgl. Cullis 1977, Alberts et al. 1983 und pp. 449/450).
Wir können daher feststellen, dass zahlreiche neuere Befunde den Ansatz Lima-de-Farias zum Thema Chromosomenfeld und seine Kritik der neodarwinistischen Chromosomen-Interpretation weiter bestätigt haben.